Back to the Syllabus
Optimal Foraging Theory
Barry Sinervo©1997
Index
Optimal Foraging and Adaptational Hypotheses
Optimal Food Choice
Optimal Clam Size in the Foraging Crow
Mussel Size Selection by Oyster Catchers
The Theorems of Optimal Foraging
The Marginal Value-Theorem: Patch Residence
Time
The Ideal Free Distribution: Patch Quality and
Competitors
Central Place Foraging
Perceptual Constraints on Optimal Foraging?
The Search Image
Rats in radial arms
Risk Aversion and Reward: Foraging Ecology of
Bumble Bees
Foraging in Juncos and Risk Aversion Under Energy
Limitation
Foraging & Biotic Interactions
Group Foraging
Predation Risk
Optimal Foraging and Adaptational Hypotheses
Attacks on the adaptational paradigm in the late 70's (Gould and Lewontin
1978) precipitated a flurry of activity in behavioral ecology, especially
in the field of Optimal Foraging Theory. Gould and Lewontin addressed
many weaknesses in the adaptationists paradigm, but the most bitting challenge
related to the notion that all phenotypic traits in an organism must
be the result of natural selection. Gould and Lewontin's arguments related
to what they referred to as the "Adaptationist's Programme".
Programme is a reference to a way of doing science or a metaphor for a computer
program that would describe how an Adaptationist (what some have referred
to as foaming adaptationists) carry out research. While the basic "adaptationist's
programme" as described by Gould and Lewontin (1978) is a caricature
of the actual way adaptationist work, their attacks did force the entire
field to look closely at the inferences people were making regarding the
utility of traits under study. Researchers became more sensitized to alternative
non-adaptational hypotheses. Despite these attacks, the Adaptationist's
Programme is alive and well. It is a step by step process in which:
- identify the trait that is likely to be under selection
- construct a functional argument for how that trait might lead to adaptive
value
- in the case of optimal foraging theory, assume that energy maximization
is related to fitness
- determine whether the mean value of the trait in the population is
"optimal" from the point of view of organismal design and function
(e.g., energy maximization).
- if the trait is not found to be under optimizing selection then go
back to step 2 and construct a new functional argument or come up with
another trait or perhaps other factors not considered that might
be leading to the discrepancy between the observed value of the trait and
the "optimality arguments".
The last step is how the metaphor relates to
a computer program. By looping back to the beginning if you do not get a
correct "optimal" answer, the adaptationist is sure to come up
with an adaptational explanation if they try hard enough. The other factors
not considered in the first loop of the program that could be taken
up in subsequent loops could be:
- other traits that cause a tradeoff,
- mechanisms that influence the function of the trait (e.g., hardness
of prey)
- constraints arising from time limitations, or
- biotic agents (e.g., predators or competitors) that might influence
the decision making.
All of these alternatives are still well within the realm of adaptational
hypotheses. Suffice it to say that Gould and Lewontin offered alternative,
non-adaptational hypotheses that might explain organismal traits.
Many of their arguments relate to constraints on design that arise from
development and organismal architecture. We will consider Gould and Lewontin's
arguments regarding constraint in some detail in subsequent Chapters and
I will briefly touch upon these ideas when I discuss memory limitations
and constraints on cognition during foraging. For the moment let us consider
the specific examples of optimal foraging theory and adaptationism and the
data that researchers have collected in recent years.
The assumption in the third step is
critical to optimal foraging theory as it is assumed that energy maximization
or optimization is in some way related to fitness maximization (e.g.,
stabilizing selection on the trait of interest). Much of optimal foraging
theory rests on this assumption and in some of the examples described below
it is clear that the researchers have to be careful to consider alternative
energy maximization rules.
I will begin a few examples of organisms that appear to be using optimal
decision making. The we will explore some theoretical concepts that are
useful in understanding the behavioral decisions that animals make and these
theoretical ideas will be followed with examples of animals foraging in
the wild. In the spirit of Gould and Lewontin, I will end with a discussion
of the perceptual constraints on foraging that might limit the choices that
animals make in the wild.
Index
Food Choice: Prey Size
One of the simplest issues in foraging decisions is picking the food
that leads to the highest rate of energy acquisition. How large a prey item
should an organism attempt to eat.
Rule number one:
Never eat anything bigger than your head.
Rule number two:
Eat items that lead to the highest gain in Energy/Time. (assuming that
you are maximizing energy.)
Profitability = Energy Gain/Unit of Time.
Index
Optimal Clam Size in the Foraging Crow
Richardson and Verbeek studied Crows foraging on clams in the intertidal
and noticed that they left quite a few clams behind after digging them up.
If they go to the trouble of digging them up in the first place why not
eat them?
The answer lies in handling time -- how long it would take them
to open the clam.
The choice would be to open this clam or search for a larger clam.
A simple calculation of whether:
(Gain small)/handling time < (Gain large)/(search time + handling
time)
would determine the optimal foraging rule.
Richardson and Verbeek computed a model similar to simple one presented
above and came up with a prediction for percentage clams eaten as a function
of size that is remarkably similar to that observed for the real data for
crows.
Index
Mussel Size Selection by Oyster Catchers
A similar problem is faced by Oyster catchers trying to open mussels.
Optimality model that only considers Energy maximization predicts that Oyster
Catchers should choose the largest clams that they encounter.
However, Oyster Catchers do not always choose the largest mussels but
leave them untouched.
Why leave the biggest items?
The handling time for the largest mussels leads to a dramatic drop in
profitability and Oyster Catchers should avoid them from the start.
The example of Oyster Catchers illustrates the approach of the Optimal
Foraging Ecologist. It is not that they are searching for reasons for a
lack of fit with an unsuitable model. Rather it is a stepwise process that
leads to more and more elaborate models. Why start with a complicated explanation
from the start -- choose the simplest explanation that will explain the
pattern.
Index
The Theorems of Optimal Foraging
The Marginal Value Theorem: Patch Residence
Time
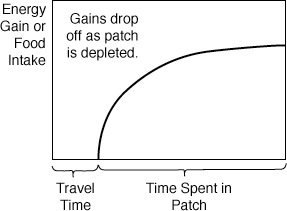 The choice about when, where, and how long to settle
or stop for a feeding bout is one of the basic decisions for an organism
that is searching for resources among widely scattered patches. One
of the simplest solutions of foraging ecology, the marginal value theorem,
is easy to derive from a graphical analysis. The marginal value theorem
yields the "giving up time" or when an organism should leave a
patch that it is exploiting. As an animal begins feeding, its energy gain
gradually begins to slow down as food becomes scarcer in the patch. It takes
longer and longer to find the next tasty little morsel, thus the curve describing
residence time in the patch and Energy Gain starts off with
a steep slope but then gradually levels off.
The choice about when, where, and how long to settle
or stop for a feeding bout is one of the basic decisions for an organism
that is searching for resources among widely scattered patches. One
of the simplest solutions of foraging ecology, the marginal value theorem,
is easy to derive from a graphical analysis. The marginal value theorem
yields the "giving up time" or when an organism should leave a
patch that it is exploiting. As an animal begins feeding, its energy gain
gradually begins to slow down as food becomes scarcer in the patch. It takes
longer and longer to find the next tasty little morsel, thus the curve describing
residence time in the patch and Energy Gain starts off with
a steep slope but then gradually levels off.
When should the animal say enough is enough and move on to find the
next patch? The crucial parameter that governs this decision is the
travel time between patches. When an animal is traveling it gains no energy.
Fundamental value to maximize: An organism should try to maximize
its net rate of energy gain (and this includes time during which
it cannot feed as it travels through a patch).
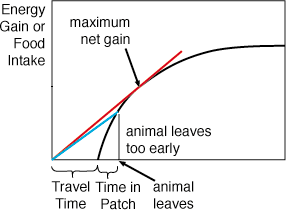 A rate of energy gain is expressed as calories gained
per unit of time. Which on the graph at the right is a straight line with
slope given by the rise over run:
A rate of energy gain is expressed as calories gained
per unit of time. Which on the graph at the right is a straight line with
slope given by the rise over run:
Energy Gain/Time.
The steepest line (gain/time) or line of greatest slope would be the
one that maximizes the rate of energy gain. Thus an, animal that leaves
too early has a shallow line (less energy per unit of time) relative to
the maximum.
There is really no benefit in staying too long as those
tasty treats are running out and the animal should move on to greener pastures.
Consequently, an animal that leaves too late also has a shallow line relative
to the line of maximum slope. The line that gives the maximum rate of energy
gain is the line that hits the gain curve at a tangent (red line).
It is the line of steepest slope, which still intersects the gain curve.
It is the maximum net gain that is possible when you factor in the travel
time between patches.
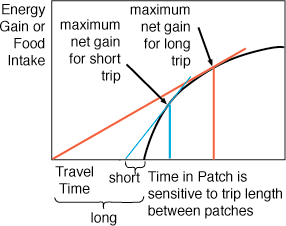 Finally, animals should also be sensitive to the length
of time it takes to travel from patch to patch. When the travel time is
short, they should leave far sooner than when the travel time between patches
is very long.
Finally, animals should also be sensitive to the length
of time it takes to travel from patch to patch. When the travel time is
short, they should leave far sooner than when the travel time between patches
is very long.
Index
The Ideal Free Distribution: Patch Quality
and Competitors
 The previous discussion ignores an consideration of variation in patch
quality, or competition from other animals. The concept of the ideal
free distribution (Fretwell and Lucas, 1972) takes both of these factors
into account in determining how animals should distribute themselves among
patches. Ideal free refers to an animal that was free to move between
patches under the ideal conditions of no constraints on movement.
In such a case, the animals should distribute themselves in the place
where the gains will be the highest. Lets consider the size of the patch
to be a measure of quality. Animals would preferentially pour into the higher
quality patch and tend to initially avoid the lower quality patches.
The previous discussion ignores an consideration of variation in patch
quality, or competition from other animals. The concept of the ideal
free distribution (Fretwell and Lucas, 1972) takes both of these factors
into account in determining how animals should distribute themselves among
patches. Ideal free refers to an animal that was free to move between
patches under the ideal conditions of no constraints on movement.
In such a case, the animals should distribute themselves in the place
where the gains will be the highest. Lets consider the size of the patch
to be a measure of quality. Animals would preferentially pour into the higher
quality patch and tend to initially avoid the lower quality patches.
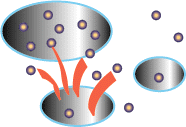 A problem arises for the individuals as more and more competitors pour
into the high quality patch. If too many competitors pour into the high
quality patch, then it would actually pay for some to move to the patch
of next highest quality as the gains from such a patch would be higher than
the first high quality patch that has now become too crowded. Animals continue
to move among patches until their sampling efforts find the highest rate
of gain. Individuals with low gain move to higher gain patches.
A problem arises for the individuals as more and more competitors pour
into the high quality patch. If too many competitors pour into the high
quality patch, then it would actually pay for some to move to the patch
of next highest quality as the gains from such a patch would be higher than
the first high quality patch that has now become too crowded. Animals continue
to move among patches until their sampling efforts find the highest rate
of gain. Individuals with low gain move to higher gain patches.
 At equilibrium, all the patches are filled up, and the number of competitors
present in each patch is proportional to the quality of the patch. Every
competitor does equally well, and it would not pay to move among patches.
The ideal free distribution is a very simple concept, but as it gets applied
to animals in the wild, it appears to have some validity. The key is to
pick the right time scale to observe animals making such ideal free decisions,
and allowing them time to come to an equilibrium.
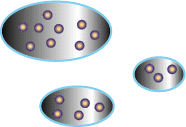
At equilibrium, all the patches are filled up, and the number of competitors
present in each patch is proportional to the quality of the patch. Every
competitor does equally well, and it would not pay to move among patches.
The ideal free distribution is a very simple concept, but as it gets applied
to animals in the wild, it appears to have some validity. The key is to
pick the right time scale to observe animals making such ideal free decisions,
and allowing them time to come to an equilibrium.
Additional references:
Sutherland, B. and Parker, G. A. 1985. in Behavioral Ecology,
Edited by R. M. Sibly and R.H. Smith, pp 255-274.
Millinski, M. 1988. Games fish play. Trends in Ecology and Evolution
3:325-330.
Index
Central Place Foraging
The concept of central place foraging is a special case of the
marginal value theorem. Animals that forage around a retreat site such as
beavers with a lodge, ground squirrels around the colony, ants around a
nest are faced with movement away from a central location to the food resources.
The question is how many items should an individual collect before returning
to the retreat, storehouse or larder. Another question arise regarding how
best to exploit the resource around the central retreat before perhaps moving
on to the next site.
The central place foraging concept is intimately related to the marginal
value theorem. In a field experiment with chipmunks, Kramer (1982) provided
the chipmunks with patches of sunflower seeds at varying distances from
their burrows. As the chipmunks stuffed their cheek pouches full, the rate
of seed collection declined in a fashion that was similar to the theoretical
gain curve. In addition, the chipmunks made
spent longer in each patch and took larger loads when they were collecting
at more distant patches to their burrow. While these trends were present,
their wasn't an exact match between the theoretical and predicated values
suggesting that other factors might have played a role in the "decision
making" of chipmunks.
Kacelnik studied central place foraging in parental starlings which have
a nest site and forage in the vicinity of the nest site for their food making
trips to collect multiple items. The question for a starling parent is how
many items to collect in its bill before returning to feed its young? Kacelnik
was able to train some birds to visit a feeder which dispensed mealworms
at an ever increasing rate to simulate the decreasing gain curve of the
marginal value theorem. In the case of parental starlings, is the parent
trying to:
- maximize its total energy gained which is given by E/time, or
- maximize energy delivered to young which also entails the energy costs
of the parent during travel as well as the energy costs for chicks or E/time
- Ep - Ec.
By manipulating the gain curve, and altering the travel time or distance
of the feeder box, Kacelnik found support for maximization rule #2. This
indicates that the answer of what is optimal depends on what the animal
might be interested in maximizing.
Index
Perceptual Constraints on Foraging?
Central place foraging assumes that animals have perfect information
regarding their environment. They do not. They have to learn about their
environment, and then as they learn, they must remember information about
their environment. These cognitive processes place limits on the kinds of
optimal choices that animals can make.
The Search Image
The first problem that an organism encounters during feeding is finding
their prey item. Prey have have evolved elaborate adaptations to be cryptic.
Classic examples of crypsis include moths on tree bark and the evolution
of crypsis during the industrialization of the United Kingdom. Around areas
with high rates of coal burning, the trees became sooty and a melanistic
form of moth increased in frequency relative to the light-colored "typical"
morph that is found on the grayish lichen in unpolluted areas. Is this a
difference that makes a difference for the moth's survival? Is there additional
natural selection on the cryptic morphs that further perfects their camouflage?
The answer to these two questions is an overwhelming -- Yes! Predators can
find a white moth on a black background more readily than a black moth (and
vice versa). Then we might ask more intricate questions regarding the abilities
of the predators.
Advocates for the idea of a search image maintain that predators
use a few cues that identify particularly cryptic prey against the background
that conceals the prey. Pietrewicz and Kamil (1979) tested aspects of the
search image hypothesis using operant conditioning on blue jays.
Under operant conditioning one tests whether subjects can learn some task,
in this prey cryptic prey discrimination, by pecking at a key in order to
receive a reward for a correctly completed task.
Their experiment had two kinds of slides presented during the Blue Jay's
discrimination task:
- a moth present on a cryptic background
- no moth present.
This experiment had the following reward structure:
- If the blue jay pecked at the key when shown a cryptic moth they received
a food reward
- If they pecked with not moth present, the did not receive food and
the jay had to wait some length of time before it could relieve the next
food item.
Experiment to test the search image:
One would predict that the Blue Jays would get better at the task (e.g.,
percentage correct) if they were presented with the same kind of moth.
Control Experiment:
However, if they were shown two different moths in a random order, they
presumably could not lock in on a search image, and they should not necessarily
get better at the discrimination task.
Results and Conclusions:
The Blue Jays got better with one type of prey suggesting that they formed
a search image. However, they did not get better when confronted with randomly
presented with two prey species with different kinds of crypsis suggesting
that they could not form a search image, or that they used the search image
formed for one species, which reduced their efficiency during episodes with
the other species.
The search image could form a short term constraint on finding prey.
The amount of time it takes to learn search images my constrain how quickly
an animal learns new ones. In addition, for organism that search for very
different prey items the search image may not be very useful at all.
Additional Reading:
Pietrewicz, A. T., and A. C. Kamil. 1979. Search image formation in the
blue jay Cyanocittata cristata. Science 204: 1332-1333.
Index
Rats in Radial Arms
Rats are thought to be central place foragers in that they move out from
a central location (typically at night) and forage along pathways to patches
with food. John P. Roche and William Timberlake looked at "The Influence
of Paths and Landmarks on the Foraging of Norway Rats (Rattus norvegicus".
They were interested in how a natural map might influence the foraging decisions
of the rats. There are advantages to following maps and thus the rat might
be tuned to follow natural maps to get around in a complex world. The maze
running ability of rats is a classic in learning theory that demonstrates
their capacity for dealing with spatial problems. Are rats constrained
in their foraging by the way they think?
The got the rats to forage in radial arms that were just "roads"
to provide structure. By manipulating the shape of the roads they could
see whether path length began to override the natural tendency of rats to
follow trackways.
Some Conclusions:
- Pathways had a larger influence on the orientation of foraging Norway
rats than vertical visual cues (beacons); they were initially more helpful
when they led to food, and more misleading when they did not lead to food,
then beacons. These differences in the salience of orientation cues disappeared
with time so that rats in different environmental conditions all foraged
with statistically indistinguishable net rate of return.
- When pathways were elongated, and thus more energetically costly to
follow, rats followed them less.
Excerpts from their discussion:
"The tendency to use paths to orient to food was influenced by the
length and location of paths. Rats in the paths-to-food treatment approached
food cups along paths a large proportion of the time. Rats in the crooked
paths, on the other hand, used paths to orient to food an intermediate proportion
of the time. Rats in the misaligned and short paths treatments infrequently
used paths to approach food cups. Rats in the crooked paths treatment displayed
significantly fewer approaches along paths, significantly more indirect
arcs, and significantly more open travel, than rats in the paths-to-food
treatment, indicating that whereas rats have a tendency to follow paths,
they are sensitive to the length of those paths relative to the distance
between food sites. Although approach of food cups from paths was low in
the short and misaligned paths treatments, rats in the short paths treatment
did approach cups from paths significantly more than rats in the misaligned
paths treatment. Rats in the short paths treatment displayed significantly
more direct arcs than rats in the other treatments with paths, perhaps because
they were less "distracted" by paths.
Some animals use the paths for more than just the shortest route to food,
but also as a means of orienting to food. However, such orientation responses
can be overridden by making it more costly to follow paths to food suggesting
that foraging efficiency will become more important and supplant the "benefits"
of the pathway approach to food.
Index
Risk Aversion and Reward: Foraging Ecology
of Bumble Bees
Leslie Real began a series of studies investigating the influence of
unpredictability in the nectar reward that foraging bumble bees encounter.
He asked a simple question that has led to some elegant questions of cognition
and its role in foraging:
Are animals sensitive to unpredictability?
Given the choice between two flowers which on average have the same reward,
but differ in variance, which will bees tend to favor.
He set up the following experimental conditions:
- One color flower, blue, initially contained 2 microliters of nectar
in every flower.
- The other flower, yellow, contained 6 microliters in every third flower
and nothing in the other two flowers.
- The flowers were placed in a large array and randomized among the color
types and rewards.
- Bees that were trained to grid were allowed to forage, and the reward
was switched between color types half way through the experiment. Blue
flowers became risky and yellow flowers became non-variable.
Bees overwhelmingly preferred the "less risky" reward flower,
even though on average they have exactly the same reward (e.g., 2 ul/flower).
Also, they switched from blue to yellow when the reward structure changed
from blue with a non-variable reward and yellow with a risky reward to blue
as a risky reward and yellow with a non-variable reward. One can even provide
the bees with more profitable but risky flowers and they still choose the
less variable flower!
71% choice for the less risky flowers
versus 50:50 predicted
Why are Bumble Bees Risk Aversive in Foraging?
Cognition and Bumble Bee Foraging
There are a number of hypotheses, however, Les Real has explored the
possibility that it may be imposed by cognitive constraints of memory on
foraging. Cognition is defined in terms of three items (Real 1993):
- perception -- a unit of information from the environment is
collected and stored in memory,
- data manipulation -- several units of information that are stored
in memory are analyzed according to computational rules built into the
nervous system,
- forming a representation of the environment -- a complete "picture"
is formed from the processing of all the information and the organism bases
its decision on the complete picture or representation of the environment.
Lets think of a simple model for memory and how it might influence foraging.
Imagine that a Bumble Bee has a specified list of items in its memory
that describe the profitability for items during bouts of foraging. It has
also been termed a memory window.
E1/T1 (last item encountered)
E2/T2 (second last item encountered)
E3/T3 (etc)....
If a bumble bee can only remember the last item upon which it foraged
(e.g., E1/T1) then a very short memory might easily explain their behavior.
Odds are 2:1 that a bumble bee will encounter 0 when feeding on the variable
reward flower. Thus, it would tend to avoid such flowers if the rule the
decision to visit the next flower color is based upon the last flower encountered.
Thus, a Bumble Bee feeding on the non-varying flower would perceive that
it was feeding on the flower with the highest reward. It would then tend
to seek out flowers with the same color.
Is a list length a constraint or an adaptation?
Real argues that such list lengths might be adaptive and not just a constraint.
If the food items are distributed randomly in the environment, then list
lengths are of no use to a forager. Feeding at one flower provides no information
about the next flower encountered. However, if the flowers are clumped in
the environment, then feeding at a flower will provide information about
nearby flowers and the foraging Bumble Bee should remember the reward and
use that information in the decision to continue feeding in the clump of
flowers or leave the clump to find a better clump. Real is collecting data
on this aspect of the model as we speak.
Additional References:
Science 1991 vol 253 980-986
Am Nat 140 suppl nov 92 s108-s145
Real and Caracao Ann Rev Ecol Syst vol 17:371-390
Index
Foraging in Juncos Under Food
Limitation
Caracao has studied the risk aversive foraging in juncos. Juncos show
the same aversion to highly variable rewards that are seen in bees. However,
if you food deprive the juncos and make them energy limited, they switch
from risk aversive foraging, to a mode that tends to select variable, but
more profitable rewards. In the example of Bumble Bee Foraging it is difficult
to reconcile risk aversion as being strictly adaptive and the arguments
regarding their choices relate to the bumble bees a built to avoid risk
in the environment (e.g., it is a product of their memory window).
Caracao demonstrated that well fed juncos are similarly risk aversive.
However if you make them energy limited to the point where they will be
in real trouble, they then will be pushed to adopt the risk seeking strategy
that has a higher pay-off.
To test this Caracao seek up two pans: the first with a non-variable
modest reward and the second pan with a variable but higher pay-off reward.
He tested the birds under conditions of being non-energy limited and
conditions of being energy deprived. The birds that were non-energy limited
chose the lower payoff pan over the higher payoff, but risky ran. When food
limitation became a factor they then opted for the higher payoff but risky
pan.
The Big Picture for Risk Aversive Behavior
Risk Aversive Behavior when not food limited is a case in which
the behaviors of the animals are not following strict optimization rules.
While researchers believe that there are still some things we do not understand
about this behavior some hypotheses relate to the possibility that memory
and cognitive constraints might limit the adaptive decision making in some
way. Such memory constraints exist at the level of the proximate constraints
that govern the processing of information and we will explore these topics
in greater detail in subsequent Chapters.
Index
Foraging and Biotic Interactions
All of the examples described above have an organism making decisions
in the absence of any real biological interactions. In the following section
we will briefly explore other factors that might influence the decision
making of organisms.
Index
Group Foraging
Why forage in groups?
If group foraging is adaptive then an organisms foraging in a group must
experience a higher payoff than if it were foraging alone. Data collected
on lions (Caracao and Wolf) illustrate the costs and benefits of group foraging.
Capture efficiency rises as a function of the number of members
in a group of lions, however, the curve levels off at large group sizes
because additional lions do not improve the odds of taking down a Thompson
gazelle.
Food availability per lion from the kill is a declining function
of group size.
Food intake per lion shows an optimal group size to be two
lions, not the 4-7 members observed in a group. Food intake takes into account
the success in foraging (capture efficiency) and the food availability.
Why do lions forage in larger than optimal groups?
This question is not yet resolved, but it is thought that it might arise
from kin selection and reasons other than foraging. Sisters may band together
in larger than expected groups because they can better able prevent males
lions from practicing infanticide (see Alcock's discussion of lion infanticide
in Chapter 1).
There may also be information transfer among members of the group
even if they are not aiding one another in taking down prey. For example,
Eric Greene (1987) demonstrated that nesting osprey in a colony pay attention
to which birds came back with successful prey items, and if they followed
such birds on successive bouts, they too were more likely to be successful
themselves.
Index
Predation Risk
For other organisms on the predated end of the spectrum, group foraging
might also alleviate the risk of predation. If an animal is solitary then
the behavior that it demonstrates with regard to foraging may not be optimal
because it must also be vigilant to predators. Many studies have demonstrated
that the animals forage in different ways with predators present than with
predators absent.
However, if animals are foraging in a group, then not every animal must
be vigilant. There are designated watchers that alert the group when predators
are near. Even if everyone still maintains some level of vigilance (e.g.,
they are not altruists) then more eyes watching lowers the overall risk
of predation than if only two eyes are watching.
Index
Back to the Syllabus