







Physics of Light, Sound, and Smells
Signal Design and Signal Detection
Motion detection and Habituation
Long-range Signature Displays and Close-range Communication
Complex Signals and Multiple Sensory Modalites
Some Case Studies of Signal Function
The classic defintion of communication involves an action on the part of one animal that alters the behavior of another (Wilson 1975). The essence of communication is the relationship between signaler and receiver. We have already discussed two kinds of signaler-receiver relationships in some detail:
In both cases of sexual selection that we covered, we discussed honesty in signal design. Implicit in the notion of honesty is the idea that signaller and receiver benefit in some way from the act of communication. For example, the female peacock who choose males on the basis of good genes, may gain material benefits for her offspring. The male that avoids conflict by advertising his supremacy in battle by a badge avoids unecessary conflict in battle and the male that is repelled by the badge avoids a costly loss.
Recall that ESS theory in male-male competition which involves the war of attrition suggests that rival males should conceal information regarding intent. However, data on animals suggests anything but concealment of intent when you look at males in the wild and during escalated conflict. Males advertise constantly, and it is thought that such advertising may be related to neighbor or individual recognition. An important component of animal communication is the idea that animal displays usually have a unique "signature" to them that allow conspecifics to identify one another. This allows animals to focus on new rivals rather than waste energy on the dear "enemies".
Transmission and reception of the signals are of paramount importance in the design of a visual system. However, these are not the only criteria for signals (Endler 1992). Guilford and Dawkins (1991) also breakdown a signals design into:
tactical components which refer to ease of transmission, detection, discrimination (e.g., from other similar signals), and ease of storage in memory; and
strategic components which refers to the signals function in a natural selection framework such as a signal that benefits the sender in some way.
Animal communication is typically thought of as being adaptive. Why else should an individual (signaller) communicate unless they are going to receive some benefit from their actions. The benefits for the signaller have already been outlined for the signals of sexual selection.
The function or adaptive value of signals has been broken down into the following classes:
While the benefits to the signaller are fairly intuitive, the benefits to the receiver can be intended or unintended. Communication with an intended receiver usually will be mutually beneficial. However, they may be many unintended receivers intercepting the signals.
Even though the signaller may receive some benefit, many animals can receive the signal. A receiver could in fact be a predatory species. Even though sexual ornaments are intended for the female, they also make the male more obvious to a predator and thus they entail costs. Advertising calls make males of many species vulnerable to a predator's detection mechanisms. Suffice it to say that there may be costs to signaller's that arise from predators that can also receive the signals.
Costs of signalling can also be found in conspecific interactions. Recall the example of alternative male strategies. Dominant males put out signals, and such signals might attract alternative morphs to the dominants. The sneakers can also manipulate signal reception mechanisms of the dominants by mimicking the displays of females. Such dishonest signalling presumably arises because of constraints on the signal reception, and processing machinery of the dominant males. One unique constraint on dominants may arise from the hormone Testosterone. Administering testosterone to chicks makes them more far sighted than near sighted. They see objects at greater distances, at the expense of close range visual acuity. Improved vision at great distance may be a tremendous advantage for a highly territorial male that has to deal with conspecific territorial males. However, far-sightedness and poor short range visual acuity could be a distinct liability in the inspection of a female mimic at close range. Perhaps testosterone results in a perceptive cost that allows sneaker males to co-opt the signal mechanisms involved in male-female interactions to their advantge and a dishonest signalling strategy can be quite successful.
There are even certain situations where animals use a dishonest signal intentionally to manipulate conspecifics. Consider male vervet monkeys that have a language consisting of a number of words for objects in the environment. Males will try to entice females into their vicinity by saying the vervet monkey word for food. The male will attempt to engage the female in copulation after this verbal sleight of hand.
The medium and sensory modalities used in the transmission of signals all seem to have a common feature -- the signals typically travel in some waveform. In the case of light, color properties of light are encode in different wavelengths of light. In addition, many visual signals that animals make also has a waveform that is generated by the body movements. In the case of sound or sonar, signals travel in waves that have different frequencies. Even olfactory cues may have a waveform associated with them as the source of the cue trails the signal somewhere downstream, the signal becomes degraded by turbulent flow, and the turbulent flow appears to impart a wavelike quality to the smell. The waves involved in olfaction could also impart information to the recipient concerning the distance that of the signaller.
Understanding vision and visual systems involves an understanding of the properties of light. While humans can see light from sun in the visible range, many animals can see light in both the infrared, and into the the ultraviolet. Some animals have exquisite color vision, others poor color vision. Others have exquisite night vision, others have poor night vision. The simplests constraints on receiver systems may involve the interactions of light with the different photo-receptors that animals have (e.g., rods versus cones). The packing of rods versus cones in the fovea of the vertebrate eye may set up a simple constraint on signal detection -- visual acuity and night vision versus color vision. Visual acuity is also a function of eye size. A larger eye casts a larger image on the retina and on a correspondingly greater number of photoreceptors (Fleishmann, 1992).
Lets consider lizards as a case study (Fleishman, 1992). Lizards have a unique eye that has two foveas. The only other vertebrate group that has two fovea's are the raptors. The central fovea of lizards is used for detection of prey or reception of signals from conspecifics at larger distances. Because lizard eyes are laterally placed, the central fovea for each eye takes in a broad field of view from one side of the animal. Given the placement of the eyes, there is very little dead space which isn't taking in an image. Each eye takes in a field of view that is 200 degrees. There is a 20 degree region of overlap at the front of the animal. This is where the temporal fovea comes into play. The temporal fovea of each eye is situated near the back of the eye (relative to the body axis) and the two temporal foveas are used in binocular vision during prey capture. Just before lunging (in anoles) or in the final dive (in raptors), the temporal fovea fixates the prey in the binocular field of vision, and this enhances depth perception.
At a more detailed level, we must also understand a little bit about the photopigments used in detecting light waves. It turns out that many vertebrates have a three pigment system. Other animals have a four pigment system. Any photopigment has an optimum wavelength that they operate at and the sensitivity of the pigment drops off around the optimum wavelength. With more and more pigments, the eye presumably has better color detection mechanisms as the coverage of the wavelengths is enhanced by the presence of more pigment optimum.
Phylogenetic history may also constraint the senses. For example, most vertebrates use a system of photoreceptors that is based on the cone. However, the source of mammalian rods may be different from rods found in other groups. Mammals are thought to have evolved from a small nocturnal ancestor that would have need excellent acuity under the difficult conditions of night. Geckos which are also active at night have converged on the mammalian rod receptor, but there are several interesting differences which point to an independent origin to the gecko "rod".
In general signals in the visual realm involve reflection or refraction for displaying color, although there are a few animals that produce light by bioluminescent processes. In addition, animals add motion, or postural changes to communicate.
Air and water are both similar in that sound travels in a wave. Because of the nature of air and turbulence, a signal projected in air is degraded quite rapidly with distance from a source. In contrast, signals in water can travel great distances before they degrade in a significant way.
The best times for signal transmission at great distance in the air is undoubtedly during the still morning when the air is also a little more dense. The tendency for territorial birds to begin their singing in the morning is undoubtedly related to the distance that such songs travel in the morning.
Water and sonar transmission in cetacea differs in one important regard compared to comparable echo location mechanisms of bats. Both forms of sound production are used in locating food items or in navigation. However, the impedance in air is very high. What this means is that a sound wave bounces back from relatively solid objects in the air because of the dramatic difference between the densities of the wave and the object. In contrast, the density of a fluid filled body in water is very comparable to the density of waves travelling through the water, and this low impedance allows dolphins to potentially be able to resolve information regarding the structure of the object that the sonar is bouncing off in their watery environment.
Sound or sonar production is a very active process, and we will look at a digital video illustrating the syrinx of the bird and song production.
The concept of input matching is central to understanding how receivers collect information. If certain colors are important from the point of view of signal origin and reception then the visual system may be fine-tuned such that only a small signal needs to be used to affect the detection mechanisms of the receiver. The spectra of light produced by the signaller may be match the optimum detection areas of the receiver.
For example, stickleback males put out a bright red signal to consepecific males and females. The visual system of the sticklebacks is fine tuned such that the emission spectra (in water) closely matches the sensitivity of the visual system of a conspecific stickleback.
Input matching may also found in the auditory system. For examples, certain calls may maximally stimulate the nerves that project from the ear.
The motion detector in most vertebrates arises from an integration of the visual system and the optic tectum or in mammals the superior colliculi. Most of the retinal ganglion cells which are hooked up to photoreceptors in the eye also project to the optic tectum. Neural output from the optic tectum is very sensitive to novel visual stimuli. The central part of the visual field and the visual neurons that project back into the optic tectum are motion sensitive to receptive fields of 0.5 degrees. In contrast the most peripheral visual fields are only sensitive to motions in excess of 40 degrees. Specific cells in the visual field are even tuned to different velocities as the respond best to a narrow range of angular velocities.
There is a reflex in most vertebrates that is sensitive to the motion detection mechanisms described above. When an object is detected in the periphery, particularly moving objects, the eye shifts such that the image becomes fixed on the central fovea -- the visual grasp reflex. Habituation mechanisms are used to shut out objects that are moving with a wavelike motion such as blowing branches. However, the eye of lizards is very sensitive to square wave patterns that lizards make during pushups and head-bobs. Lizards are very responsive to square waves and are not very responsive to sine wave motion.
Finally habituation is a key concept in motion detection (and indeed in other sensory modalities). If the motion is period and the motion cycles through 2-3 cycles, the response of the motion detector neurons becomes habituated and the response is turned off. Such habituation mechanisms allow the animal to shut out noise such as the period movements found in vegetation blowing in the wind. Similar habituation mechanisms are found in the auditory system for removing white noise (noise of random frequency and amplitude).
Lizards use the square wave signal periodically during their territorial patrols and the signature display that they use is of very large amplitude. The signature display of many lizards that have been studies include unique individual components to them much like bird song or whale song has an individuality to it. In addition, the signature display in most organisms is usually designed to function as a species recognition signal so it is also very conserved in other key respects. Finally the Signature display is designed to be received at fairly large distances. In lizards that might entail a few territory diameters. Bird song signatures are meant to be heard over several territory diameters. The nature of the ocean as a medium for the transport of signals allows whales to transmit their signature displays over hundreds of kilometers.
Signature displays are meant to get an individuals attention. Once the other animal is listening or watching the animals will typically move in to closer range, and begin signalling with smaller amplitude or more quite signals. In the case of lizards, two males that are at close range use a much smaller amplitude "challenge display". In the case of a male and female making contact, the courtship song at close range is much more quiet than the territorial signature display that is meant for long range reception.
Many signals that animals make that are important do not rely on a single sensory modality (e.g., vision vs sound), but can use a number of modalities. For example, lekking in golden collared manakins involves:
Large herbivores give a "pursuit invitation display" that involves two visual cues. When they detect a distant predator, deer (and gazelles) lift their tail to reveal a white patch of fur and they also move away with a stiff legged stotting gait. The signal is presumably used to signal the predator a message of which would have the following effect (e.g., in anthropomorphic human terms): "you have been detected and the jig is up -- pursuit is futile."
Animal signals are tuned to the environmental conditions. Why make a big or "noisy" signal when a small signal will suffice. For example, squirrels found in woodland habitats use a very large "tail flagging" motion, where the tail is moved from the back to the nearly the ground. In contrast, a species found in rocky habitats use a short jerky tail flick. Similarly sticklebacks in dense weeds court females with a jump dance to attract the attention of the female, whereas sticklebacks found in barren environments do not appear to use the jump dance at all (Fleishman, 1992).
The lighting conditions that animals are found in also affect signal design structure (Endler 1992). The most striking differences would occur with:
A recent theory (Endler, 1992, Ryan 1990) of the 1980's is that there exists a sensory bias in the nervous/sensory receptors of females that pre-disposes them to pick some male traits, not because they perceive them as sexy per se, but because they are "attracted to them", probably for reasons other than mate choice.
Certain stimuli (e.g., colors, shapes, movement) may be useful in other contexts (e.g., feeding and foraging) and the nervous system of females (and males) are honed by natural selection to be efficient at picking out food items from a world that is overly rich in extraneous stimuli.
In a sense, these parts of the nervous system/sensory system may be co-opted by sexual selection and males that show a trait that triggers a hightened response in females may have an advantage. A males signal may become fine-tuned such that it maximally stimulates the female sensory system.
<See Xiphophorus tape by Molly Morris>
Basalo looked a genus of Sword-tailed fish, Xiphoporus, which have elongate swords. A phylogeny of Xiphophorus indicates that most recent members of the "clade" have swords. One member of the genus, the most "ancestral" type lacks a sword. Females mate with males without swords.
Basalo asked whether females from this ancestral species preferred males of their own species which lack a sword, or males of their own species with swords tied on. The overwhelming choice was for males that had a Sword!!! She interpreted these results to imply that there existed an "ancestral" bias, for swordedness in these fish, that in turn led to a Runaway Process.
Any colony nesting animal with parental care faces a dilemma. When the parent(s) leave to forage, they must be able to recognize their offspring when they return. This might be simple in an animal that can navigate back to a fixed nest where the locations of the nests does not change. However, many animals such as bats, birds, and marine mammals are found in large colonies without very distinct boundaries.
Selection strongly favors the evolution of individual recognition between the parent and the offspring and a simple video example for southern elephant seals on Kangaroo Island (courtesy of Dan Costa) illustrates this quite nicely.
Whales and dolphins are constantly projecting noises, through their sonar clicks and whistles. Much of the sonar is used during navigation. The sonar pulses are reflected back from objects in the environment, and the whales either steer clear of these objects, or if the object is prey at close range they move in to intercept.
Can such navigational signals also have an indirect role (or direct) in communicating information about the environment to other whales. Clarke and Ellison (unpublished) tracked whales as they approached a large ice flow. The lead whales or earlier arriving whales detected the ice flow at fairly close range and moved around the ice flow accordingly. However, later arriving whales appeared to anticipate the ice flow and had a much straighter trajectory around the ice than the earlier whales suggesting that they had used information from the previous whales encounters to adjust their paths accordingly.
Was the receiver obtaining unintended information. The signaller's produced a sound (lead whales) and it had an effect on the receivers behavior (trailing whales). Under Wilson's broad definition it would be considered communication.
For years the actual function of whale song has been somewhat enigmatic. Recently, playback experiments in hump back whales reveal some interesting functions of the haunting song. Most cetacea appear to have a very species specific song pattern that also appears to double as a signature display. This song is fairly simple.
However, humpbacks and other whales enter into long exchanges and the songs used in these exchanges is much more complex. In addition, it is clear that individuals modify their songs year after year, and the population as a whole tends to show a similar "cultural" transmission to song that is seen in birds. Songs within a localized population appear to be more similar than more distant populations.
Tyack presents compelling evidence that these songs are used in male-male competition. Males will come up to speakers that are broadcasting an "intruder" male's song. The "resident" (for lack of a better term in whales) will then broadcast its own song which may in fact represent a challenge display. If males use such songs in male-male disputes, it is likely that females use such songs in female choice as well though critical experiments remain to be designed.
While the typical notion is that males are directing the course of courtship, female cowbirds have a number of options open to them in response to a male's song. Some of these responses prevent the male from being able to establish a vocal history with the female. For example, a female can simply hide herself away from the male. In aviary observations, while males are usually prominent and easily seen, sometimes individual females cannot be found. Go to the overview of the cowbird dating game for more detail on mating strategies.
|
" FLY AWAY" Often a female will fly away as soon as a male sings to her. This effectively prevents him from singing another song at this location. Flying away from the male sometimes leads to him chasing her, so singing might be resumed in a new location. Quicktime format (millions of colors 318 K). |
|
" FLY AWAY BEFORE SONG" A female can anticipate the male's song by his approach, and she will frequently fly away before he can even begin to sing. In this clip the female leaves as soon as the male moves into position to sing to her.Quicktime format (millions of colors 572K). |
|
"IGNORE" The most frequent response to song by the female is a lack of any overt attention. Her behavior continues as if the male had not sung. Sometimes a male will follow a female for several minutes while singing song after song to her, and she simply ignores him. Quicktime format (millions of colors 1.7 mb). |
|
"LUNGE" This clipshows the female lunging at the male as he sings. The lunge certainly appears to the human eye to be an attempt to chase away the singing male. However, the male seldom leaves after a lunge, and will frequently stay and sing another song, making the function of the lunge unclear. In this clip, the male appears to fly away following the lunge, however, he only moved further down the branch, out of our view. Quicktime format (millions of colors about 1.2 mb. This video is recorded in half-time slow motion). |
|
"RATTLE" The female has a rattling vocalization that she sometimes uses during a song. It's function has not been determined. The rattle may be an attempt to mask the sound of the song, or it may be a signal indicating interest. It is a general response that can also be accompanied by a lunge or by flight, and is sometimes even heard during a sequence of songs that leads to a copulation. (millions of colors about 826 K. This video plays at 25 frames per second in order to capture the speed of the rattle). |
|
"COPULATION" This clip shows the culmination of what is usually several days of constant singing: a successful copulation. A male might sing several hundred songs to an individual female before she will adopt the copulatory posture and mate with the male. Quicktime format (millions of colors about 1.7 mb). |
|
"LONGER COPULATION" This is a longer sequence showing a copulation with several songs leading up to it. (millions of colors 4.3 mb; this will take longer to download). |
|
" MALE/ MALE SONG." "MALE/MALE SONG"Males also sing to one another, as shown in this clip. Male/ male interactions are thought to function in establishing social order. They are also observed by females, and perhaps used to help assess the worth of an individual male. |
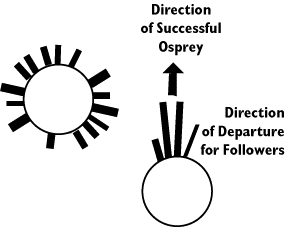 On the left side of the figure you see the histograms that describe
the directions that osprey take when no bird has returned with prey. On
the right you see that birds that take off in response to a successful forager's
return head off in the direction of the successful forager. These followers
find food in 1/3 the time compared to naive osprey. Eric Greene has also
described a dance that seems very similar to the orientation dance of bees.
On the left side of the figure you see the histograms that describe
the directions that osprey take when no bird has returned with prey. On
the right you see that birds that take off in response to a successful forager's
return head off in the direction of the successful forager. These followers
find food in 1/3 the time compared to naive osprey. Eric Greene has also
described a dance that seems very similar to the orientation dance of bees.
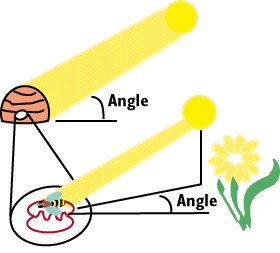 Recall that bees go out on foraging flights, find food, then fly back
to the hive to communicate the location of the food source to the other
workers.
Recall that bees go out on foraging flights, find food, then fly back
to the hive to communicate the location of the food source to the other
workers.
The bees use a waggle dance to indicate the direction of the nectar source.
The sun provides a very constant reference in the sky for the bees to
direct other workers. The bee ends its circuit around the loop with a waggle,
and the distinctive waggle part of the dance lines up with a vector that
leads directly to the nectar source. The distance from the hive is proportional
to the number of waggles in the dance.
Ground squirrels are notorious for their altruistic alarm calls. Why on earth should a Ground squirrel scream at the top of their lungs. This advertises their location to the predator. It also appears to alert other memebers of the colony, and the colony mates scatter in response to the alarm call. Ground squirrels also have distinctive call patterns for:
caller might be alerting kin, even though it is raising its own risk of mortality
Sherman eliminated A.1-3.
But found support for A. 4 and B. (see Alcock for details).
Vervet monkeys show an even greater number of prey types in that they recognize cats, large raptors, and large snakes. Alarm communication is also found in the more primitive lemurs which likewise recognize and use alarms for birds and terrestrial predators.