Back to the Syllabus
Speciation Mechanisms
Barry Sinervo©1997
Index
Biological Isolating Mechanisms
Modes of Speciation Classified by Geography
Allopatric Speciation
Ring Species and Species Recognition in Gulls
Parapatric Speciation and Allo-Parapatric Speciation
Sympatric Speciation
Conditions for Sympatric Speciation -- Assortative
mating
Multiple-Niche Polymorphism in Insects and Host-Plant
Preference
Runaway Process, Sexual Selection, and Speciation
Speciation Mechanisms
Biological Isolating Mechanisms
"Species are groups of actually or potentially interbreeding
populations, which are reproductively isolated from other such groups."
Mayr 1942
Ernst Mayr's formulation of the biological species concept clarified
a vexing question for evolutionary biologists: What is a species? However,
how species arise by the process of natural selection remains a source of
great controversy. The problem is that the process of speciation takes a
very long time to complete, and it occurs on a very large geographic scale.
In addition to Mayr's definition the following concepts are quite useful
in delimiting species.
Hybrid sterility definitely separates two species, although it
is not the sole criterion
Reproductive isolation could however be due to behavioral or morphological
blocks to mating (i.e., hybrids are possible but unlikely given such blocks).
Barriers are not merely geographic, but have a biological origin
and arise during the process of speciation.
Isolating mechanisms:
A) Premating isolating mechanisms -- prevent union of gametes
-> zygote
- mates do not meet (seasonal or habitat isolation)
- mates meet but do not mate (ethological or behavioral isolation)
- mates meet but no sperm transfer (mechanical isolation)
B) Post-mating isolating mechanisms -- varying degrees of hybrid
sterility
- sperm transfered but dies before fertilization
- zygote dies
- zygote produces an F1 adult that has reduced viability (survival)
- hybrid is viable, but partially or completely sterile (fecundity) or
the F2 is deficient.
Species might simultaneously possess both premating and post-mating isolating
mechanisms.
As we will see below, the biological species concept begins to break
down in areas or at times of speciation or incipient speciation (semispecies).
For example at hybrid zones, the somewhat differentiated populations
are in the process of speciation. From the point of view of individual selection
and the fitness of an organism in such a hybrid zone, the fundamental question
regarding animal behavior and speciation is:
Which of the two mating isolation mechanisms is the most effective in
reducing the risk of producing low fitness offspring? Which mechanisms
is most efficient in terms of time and lost reproductive opportunity?
An organism that mates with a different "semispecies" might
produce low viability offspring (in any of the degrees listed above) and
such a beast has lower fitness compared to an organism that discriminates
against such semispecies and mates with members of its own semispecies (e.g.,
posseses isolating mechanism A.1, A.2, or A.3 or perhaps all of the above
discrimination mechanisms). Such behavioral descrimination ability should
evolve rapidly in the area of incipient speciation, under the right conditions.
What are the right conditions? This is a very difficult question to answer.
It requires an understanding of the field of population genetics and the
crucial parameter is how much migration occurs from areas where each
semispecies exists in its pure form. A second parameter, has to do with
the degree of genetic similarity between the two pure semispecies.
Migration is the movement of animals between what are referred
to as populations of interbreeding animals. Rates of migration are usually
expressed as the proportion of individuals in a population that disperse
from one population to the next each generation. If migration between the
region of contact between the two semispecies and the rest of the semispecies
ranges is too high, then the two semispecies will become homogenized into
one species even if such interbreeding results in lower fitness because
of hybrid unfitness. If migration is low enough, then the two pure semispecies
will eventually become differentiated into two species owing to the evolution
of isolating mechanisms. Evolution of such isolating mechanisms is contigent
upon the degree of genetic similarity between the two semispecies.
Large differences in genetic similarity between semispecies are
more likely to produce unfit hybrids than small differences in similarity.
If the two semispecies have many genetic loci that differ in the kind of
allele that they possess, then those alleles might not work very well together,
and more importantly the way two different genetic loci work together may
begin to breakdown. This is referred to as genome-wide genetic interactions.
The correct operation of one genetic locus (lets say an enzyme) may require
a particular form of the enzyme that is produced by a different locus (e.g.,
epistasis). These enzymes are most likely to be compatible if they come
from the same semispecies. If they are from different semispecies, they
may not work at all and the hybrid individual may die. Imagine such genetic
interactions occuring at many different loci (e.g., 4000 loci in Drosophila
or perhaps 100,000 loci in humans). The probability of a hybrid being more
unfit has to do with how many loci in each semispecies are homozygous for
different alleles. Hybrid breakdown occurs when many of these
loci are fixed for different alleles in the two subspecies and the production
of offspring by the hybrids is not possible because and F1 X F1 produces
too many incompatible gene by gene combinations.
Genetic similarity drops as we change from the different levels
of differentiation, going up the heirarchy:
Populations - Subspecies - Semispecies - Sibling species - non-sibling
species - species
By the time species are compared there is generally NO GENE FLOW.
As genetic similarity drops, the differences increase and the possibility
of hybrid breakdown increases. A species is thought to have a coadapted
genome in which many genes are finely tuned by the proper interactions
with other genes.
Clearly genetic differentiation is occuring during speciation but it
is not enough to define a species. It can vary from organism to organism
and from mode of speciation to the next. In practice, even the criterion
of NO GENE FLOW between species is not the defining feature of species.
Index
Modes of Speciation Classified by Geography
To round out our discussion of speciation, the ways of categorizing speciation
as a process is discussed below. This is a review of material from a course
on evolution. The geographic subdivision acts to decrease gene flow and
promote the process of speciation.
- Allopatric Speciation -- speciation occurs in geographic isolation,
- Parapatric Speciation -- speciation in adjacent populations
with gene flow,
- Allo-Parapatric Speciation -- populations initially separated
(allo-) but then secondarily come into contact with subsequent parapatric
speciation,
- Sympatric Speciation -- speciation within a panmictic population.
These modes of speciation relate to the degree of geographical subdivision
which ranges from:
Extreme <--> None
for the modes:
Allopatric -- Allo-parapatric -- Parapatric -- Sympatric
Patterns of variation across the species range is at the heart
of the matter and the presence of clines in morphological or behavioral
traits is used as evidence for one mode of speciation over another. A cline
is any systematic change in a trait across the range of a species. Clines
can be abrupt or gradual.
CLINES:
- Presence no cline. -- no contact, species have no gene flow
due to isolation -- Allopatric Speciation Rules.
- Shape of the Cline. To what degree is the variation clinal (e.g.,
at a hybrid zone). Clines can be gradual along a transect or very abrupt
(within a few 100 m whereas a species is spread over 1000 km). Is
the cline maintained by persistent selection? If yes, then the example,
might be a candidate for parapatric speciation.
- Origin of the cline or hybrid zone. Is it due to secondary contact
with subsequent weak gene flow across the edge. Evidence for secondary
contact can include concordant variation in many characters on either side
of the cline, if so the system may have evolved by allo-parapatric speciation.
- No cline but sympatric -- sympatric speciation in situ due to
temporal, ecological, or behavioral segregation. Clines are not an issue.
Index
Allopatric Speciation
Allopatric speciation occurs under the following conditions:
- isolation of a colony (e.g., island and mainland)
- division of range by extrinsic barrier (e.g., mountain range) or extinction
of populations in the intermediate parts of the range.
- isolation by geographical distance.
All of these factors severely restrict gene flow and allow populations
of a species on one side of the range to begin to differentiate from species
on the other side of the range. Given enough time, the genetic differences
became large enough to warrant calling the "races" on one side
or the other a different species.
Ring Species and Species Recognition in Gulls
 Ring Species arise from a peculiar form of allopatric speciation
that takes place when the center of a species range is effectively unoccupied.
Adjacent races all around the ring will interbreed, however the races at
the termini are so divergent, that there is no interbreeding, and they can
exist in sympatry without interbreeding.
Ring Species arise from a peculiar form of allopatric speciation
that takes place when the center of a species range is effectively unoccupied.
Adjacent races all around the ring will interbreed, however the races at
the termini are so divergent, that there is no interbreeding, and they can
exist in sympatry without interbreeding.
The species range of Sea Gulls in the Arctic Ocean represents a classic
example of a ring species. The polar ice cap limits the species range of
Sea Gulls to a ring that circumnavigates the globel In addition, work by
Smith demonstrates the key reproductive isolating mechanism at the "ends"
of the ring.
There are a number of races of Sea Gulls as one traverses around the
globe. Races from America freely interbreed with races from Europe. Races
from Siberia freely interbreed with races from America. Races from Siberia
freely interbreed with races from the Caucauses. However Centrial European
races do not interbreed with those from Western Europe. All along the ring
there is gene flow, but where the two ends of the ring meet in Europe there
is not gene flow.
Smith investigated the signal that is used to isolate the two species
into Larus hyperboreus, the glaucous gull and Larus thayeri,
Thayer's Gull. The glaucous gull has a yellowish eye ring, and Thayer's
Gull has a purple eye ring. Smith painted the eye rings of Thayer's males
yellow, and this allowed Thayer males to successful pair with glaucous females.
However, the males would not mate with the glaucous females until Smith
painted the glaucous females purple. Such paint experiments are a powerful
means of demonstrating the meaning behind signals and the role in species
recognition.
Index
Parapatric Speciation and Allo-Parapatric
Speciation
In order for a species to arise by parapatric speciation, there
must usually be a very strong change in the environment. Natural selection
on one side of the environmental transition favors one set of traits compared
to the traits favored on the other side. The existence of clines as evidence
for parapatric speciation is generally not considered very good evidence.
The ring species of gulls forms a cline in that there are small changes
along the length of the ring.
However, if selection is strong enough it leads to strong enough disruptive
selection at the abrupt transition that parapatric speciation occurs.
Animals that do interbreed (e.g., mate with a different type) at such
hybrid zones tend to show underdominance in fitness, that is their
progeny the heterozygotes (hybrids) have lower fitness. Another way to think
of this pattern is to view it as disruptive selection against individuals
heterozygous for a large number of loci. The genes in the heterozygous condition
have lower fitness than homozygous configurations because of the strong
environmental differences on either side of the contact zone.
Introgressive hybridization -- If there is any viability of the
hybrids, alleles may leak across and introgress across the zone. Some alleles
may spread quite far across the zone whereas others will be quite abrupt
and coincide with the hybrid zone.
Big Question: Can we distinguish between primary and secondary
contact? In principle, we cannot which is the case and that is why the models
of speciation are so fiercely debated.
Index
Sympatric speciation
Conditions for Sympatric Speciation -- Assortative
Mating
Lets re-visit an old system -- the beak morphs of African finches (Smith).
Let us speculate about what might happen to Pyrenestes in the not
too distant future.
If S (small-billed) birds are much more successful in feeding on small
seeds and L (large-billed) birds are much more successful in feeding on
large seeds, what if S birds tend to mate with other S birds, and L birds
tend to mate with other L birds? Why might the birds mate assortatively?
Let us look at the data. Tom Smith collected data on assortative mating.
He observed the following frequencies of matings Sm X Sf = 34, Sm X Lf =
14, Lm X Sf = 14, Lm X Lf = 6. Lets do a few simple computations to see
if the birds are breeding randomly or are the two morphs associating with
similar morphotypes.
| |
female of the the pair |
S |
L |
| male in the pair |
S |
34 |
14 |
L |
14 |
6 |
What would we expect by chance? We need to compute the following items.
What is the frequency of small billed males?
(34+14)/(34+14+14+6) = 48/68 = 0.71
What is the frequency of small billed females?
(34+14)/(34+14+14+6) = 48/68 = 0.71
What is the frequency of large billed males?
(14+6)/(34+14+14+6) = 22/68 = 0.29
What is the frequency of large billed females?
(14+6)/(34+14+14+6) = 22/68 = 0.29
What is the probability that a small billed male is randomly paired with
a small billed female?
0.71 * 0.71 = 0.54
and we would expect to see
68 * 0.54 = 34.3.
By the same logic we can compute our random expectations for the other
three kinds of matings to derive an expected number of matings if the birds
were randomly mating.
| Observed matings |
Expected by Random Mating |
| |
female of the the pair |
S |
L |
| male in the pair |
S |
34 |
14 |
L |
14 |
6 |
|
| |
female of the the pair |
S |
L |
| male in the pair |
S |
34.3 |
14 |
L |
14 |
5.7 |
|
Given the answer from this test, how likely is sympatric speciation in
these birds? Not very likely!
Lets contrast this with pure assortative mating in which the birds breed
with the same type. We should see something like the following data for
sympatric speciation to be likely.
| |
female of the the pair |
S |
L |
| male in the pair |
S |
48 |
0 |
L |
0 |
20 |
Index
Multiple-Niche Polymorphism in Insects and
Host-Plant Preference
Sympatric speciation is the origin of two new species with the two forms
in the same place. Most other theories involving a gradual change in sympatry
are controversial except for the following case that involves plants and
insects. It is thought that the following mechanism may be why there are
more insect species of insects on the planet than any other organism. If
so, then this mechanism is truly important from the point of view of animal
behavior and diversity. Part of the reason why it is so prevalent is because
insects can evolve detoxifying specificity towards their host plants in
a specific enzyme -- P450 (the details of which need not concern us). However,
the acquisition of such specificity should lead to rapid acquisition of
host plant preferences -- a behavioral trait.
One class of models maintains that there arise multiple-niche polymorphisms
(heterozygotes inferior). Assortative mating among homozygotes is a
key component of this theory. This minimally requires two loci to evolve
simulatneously, and thus we establish strong linkage disequilibrium for
the niche polymorphism loci and the assortative mating loci. More loci makes
it unlikely to occur.
One classic and well accepted form of sympatric speciation involving
animal behavior has to do with host plant choice by insects. Many host plants
of insects produce very nasty chemicals in repsonse to being eaten. The
chemicals are meant to deter the herbivores, thus in order to continue feeding
on the plants, the insects must develop mutations that allow it to detoxify
the chemicals. Because different host species produce very different toxins,
the mutations that allow an insect to detoxify one host-plants arsenal of
toxins do not work with another. Here are the key issues involved in this
mode of speciation.
An insect must evolve the detoxifying mutations, and also evolve
host-plant preferences that allow it to become very choosy about
the kind of plants it is feeding on.
The mutations do not have to occur simultaneously, but the detoxifying
mutation probably arises first and the behavioral preference evolves
as a refinement to the detoxifying phenotye because individuals that match
their own detoxifying abilities with the proper host plant would be strongly
favored by natural selection.
In general, two such mutations are required, because it is difficult
to imagine getting a single mutation that has a pleiotropic effect on detoxifying
ability and host-plant preference.
The two mutations should also arise on the same chromosome in close proximity
to one another. This causes the behavior for host-plant specificity to be
tightly correlated with the detoxifying genes. The linkage between the
two loci forms a genetic correlation between the behavioral and physiological
traits. An insect that ends up with the wrong allele for detoxifying matched
up with the wrong preference is in trouble (recall the problem for stripped
and spotted snakes). By having the mutations on the same chromosome, a super
gene for the detoxifying ability and host-plant preference is produced.
It is possible for the genome to become rearranged after the mutations arise.
This would require a third chromosomal rearrangement mutation which makes
it less likely.
Technically, the incipient insect species which feeds on a new host plant
species is found in the same area as the ancestral species, but by virtue
of different host plant preferences and mating microhabitats, they no longer
exchange genes. The easiest way for many insects to find suitable hosts
for their own progeny would be to oviposit their eggs on the host plant.
If you oviposit on your host plant, chances are mating would also occur
on the plant prior to oviposition.
This speciation process not only leads to adaptive plant choice, but
because other members of the insects own incipient species also choose the
same plants, the insects will mate with like members of their incipient
species and not with other members, and reproductive isolation is achieved.
Index
Runaway Process, Sexual Selection, and Speciation
Recall the elements of Fisherian
runaway process and sexual selection. The essence of the theory is that
a mutation for female choosiness arises in a population and the females
opt for males with some kind of elaborate trait. If Fisherian runaway process
is to play a role in speciation mechanisms, then it must be related to the
process by which reproductive isolation occurs.
How do the female descrimination traits such as that seen in Gulls evolve.
What about plumage variation seen in other bird species? How is it that
lizards have evolved such markedly divergent head bobs.
The theory for runaway process as independently solved by Mark Kirkpatrick
and Russel Lande has a very interesting second phase.
In the first phase we saw that:
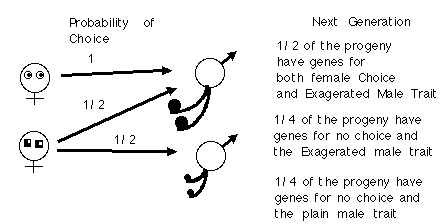
Fisherian runaway will lead to ever increasing escalation in the male
trait up to the point that natural selection begins to strongly act against
the handicap that the male trait imposes on the male. Once this point is
reached, the second phase of Fisherian runaway sexual selection takes hold.
The populations will change in frequency by the process of what is refered
to as genetic drift. Genetic drift is simply the random processes
that cause small fluctuations in the frequency of an allele simply because
of limited population size. By chance alone, the frequency of alleles (for
choosiness and the male trait) will fluctuate up and down in frequency.
In small populations the effect of genetic is expected to be quite great
compared to large populations. How small is small? Well generally populations
that are less than about 100 individuals. What might happen is that certain
alleles for choosiness might go to fixation in a small population, or perhaps
genes for the male trait.
Up to this point we have considered runaway selection and its operation
on a single male trait and a single mutation for choosiness. However, as
discussed above sexual selection operates on the whole
organism and females might choose on the basis of a number of male traits.
For example, females might choose on the basis of both tail length as well
as plumage coloration.
Now imagine a number of distinct sub-populations which are experiencing
genetic drift and going to fixation. Because genetic drift is an entirely
random process we would expect that each sub-population would fix for different
choosiness or male traits. When this occurs, we have evolved strong assortative
mating mechanisms that would lead to isolation.
Another scenario for such random effects is that the mutations
that lead to choosiness or the male trait might differ in each sub-population.
Fisherian runaway selection might lead to rapid evolution in each population
according to such initial differences. And the populations would be expected
to diverge rapidly in their mating preferences.
The end result of either sub-mechanism of runaway is reproductive
isolation by mating preferences -- the key ingredient for speciation.
Index
Back to the Syllabus