



male brood pouch
copulation
full brood pouch
birth
fry and father
The Origin of Male Ornaments and Female Choice
Parental Investment: Why Females Choose and Males Display?
Models of Mate Choice and Experimental Tests
How Female Choice and the Male Trait Come to Be Correlated
Evidence for Genetic Correlations Between Female Choice and the Male Trait
Good Genes or Indicator Models
The Origin of Female Choices and Male Traits
Sensory Bias or Sensory Exploitation
Charles Darwin was the first to realize that sexual selection by female choice or male-male competition was likely to lead to the evolution of elaborate structures which reduce survival. In the case of male-male competition, the direct costs of such structures arise from the long and protracted battles and the possession of a weapon per se. While lethal fighting is rare in the animal kingdom it still does occaissionally occur in many species. Moreover, even if battle is not lethal, the war of attrition can leave both combatants in a weakened physiological state and precipitate their death. In addition, the physiological mechanisms that produce such structures have indirect costs which reduce survival. For example, the development of many secondary sexual traits in vertebrates requires the production of Testosterone. If the production or development of a larger structure requires more testoterone, then such hormones might lead to survival costs because of a pleiotropic relationship between survival and the sexually selected traits governed by testosterone production.
The reduction in survival arising from male-male competition is not all that enigmatic given that elimination of competition from rivals is the primary goal of male-male competition.
However, why should female choice promote the evolution of ornaments in males that carry heavy survival costs?
We will explore the role of female choice in models of sexual selection to address this question.
The simplest answer to such question is that male ornaments are used to attract females and the ornaments are also used in male-male competition. A female that is watching a male contest is likely to choose the victor, and her choice may be directly tied to the outcome of male-male competition. In most animals many structures are used in both female choice and male-male competition. Most studies of female choice tend to focus on structures that have only an "attractive" role. Many researchers assume that because a structure is not used directly in battle, that structure is used primarily by females to choose a mate. Recall from the chapter on Male Strategies, that Badges of Status might evolve as an information signal which announces the males physiological vigor to rivals. The male does not use his throat color or bright plumage in battle, but the male might also use it as an intimidation tactic and avoid costly battle. Keep in mind that issues of female choice are very difficult to disentangle from male-male competition.
The role of the female choice in promoting sexual selection is largely tied to the the observation that females invest more heavily in reproduction and thus this leads to females being more limiting. A female that is taking care of young or gathering energy to provision eggs is not interested in mate choice after she copulates with the male. Higher parental investment by females was advocated by Trivers (1972) as the primary mechanism driving strong sexual selection on male traits and for female choice. Whereas the females may invest in prolonged care in young, the male is free to move on and mate with other females or compete with other males for access to females. Thus, females should be quite choosy about their mate because males are present in abundance. The role of parental investment in mate choice has been powerfully tested in animals with sex-role reversal where the male is primarily responsible for parental care.
Consider the stickleback in which the male builds a nest and guards the young after they hatch. The male is the primary care giver, but in sticklebacks, the female is still the choosier sex, and females choose males on the basis of the bright red color on the male belly.
Why are females still choosing male sticklebacks based on ornaments?
The nest of a male stickleback is not a limiting resource. A male stickleback builds a nest but nest size is not all that limiting. A male stickleback can still attract many females to his nest and thus, sexually selected traits might be strongly favored. In addition, a male that is successful and vigorous in defending his nest from rivals may also become a good parent in that he must aggressively defend his nest and offspring from rivals and predators.
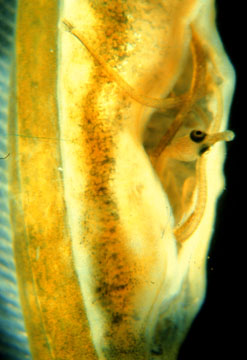
The conditions for the evolution of male choice are not quite met in sticklebacks because males are not the limiting sex, female egg production might still be limiting for males. The idea of operational sex ratio (Emlen and Oring) explains why male stickleback are under strong sexual selection and females still choose males on the basis of their bright red coloration. The reproductive rate of the population is still limited by females which must yolk up additional eggs for subsequent clutches. At any point during the breeding season, the number of males that are available for mating far exceeds the number of available females. Many females are in the process of yolking up eggs, whereas all nest guarding males are still available to fertilize a female's clutch of eggs. The operational sex ratio is still male biased in that there are fewer females available for fertilization even if there are equal numbers of males and females that are reproductive. Conditions in the stickleback mating system still favor choosy females. Let us consider a close relative of the stickleback which has evolved a higher level of male care -- a brood pouch.
Sticklebacks, pipefish, and seahorses belong to the same family of fish and this group is noteworthy in the animal kingdom for the evolution of highly-advanced male care. The care in sticklebacks is limited to nest defense, and this is a relatively common occurrance in the animal kingdom (e.g., many families of fish, birds, mammals, insects, etc have males that guard the nest).
However, pipefish and seahorse males develop an elaborate brood pouch during the reproductive season. In some species, the brood pouch is simply a place where the female oviposits her eggs. However, in other species, the males have evolved elaborate vascularization in the pouch and they can transfer nutrients to the eggs much like a female mammal transfers micronutrients across the placenta. The evolution of such elaborate and expensive male care in pipefish causes males to be the limiting sex and the operational sex ratio is female biased in that excess females are available for reproduction.
| |
|
|
|
|
male brood pouch |
copulation |
full brood pouch |
birth |
fry and father |
In addition, the size of the male's brood pouch is a limiting resource in pipefish, in contrast to less size limitation in the case of a male stickleback's nest. A male pipefish can only hold a limited number of embryos and perhaps he should be quite choosy about the quality of the female.
In two species of pipefish females have evolved elaborate ornaments to attract males. Males are now the choosy sex owing to females being available in abundance. Rosenqvuist assessed the importance of the female ornament in male choice by presenting females of different size to males and presenting females which varied in the degree of ornament development. Overwhelmingly, males preferred females that were larger and more ornamented, the key conditions necessary for the evolution of even more elaborate size and ornamentation in female pipefish.
 |
 |
Nerophis Ophidon male |
Nerophis Ophidon female |
The case of sex-role reversal in pipefish and seahorses represents an extreme departure from the typical system where females give care to young and males do not. Even though the males of most species do not participate in care or the acquisition of energy for young, males can provide nutrients to the female in the form of nuptial gifts. The nuptial gift provided by the male can be a prey item that is offered to the female in the case of hangflies, or even the male himself in the case of some species of blackwidow. In some blackwidow species, males will approach the female, attach their gentalia, then he will do a backflip into the females fangs. While she is eating the male, sperm transfer is completed.
The most elaborate nuptial gifts are a special spermatophore packet that contains the normal sperm packet along with a nutritious protein rich spermatophore. The male offers the female the gift, and places it on her gentalia. While she feasts on the treat, the sperm is transfered into the female. The larger the spermatophore packet, the longer the sperm transfer, and the more eggs the male will fertilize in her brood. If a male transfers too small a packet, the female might finish off the protein packet and munch away on the sperm before it is transfered. The female also benefits, because she produces additional eggs with the energy that she received in the gift.
 Males might choose females on the basis of the quality
of the male or the quality of the territory that the male secures. Let us
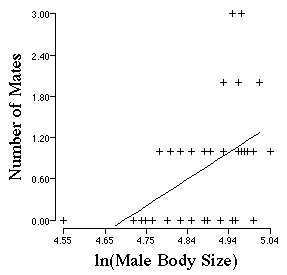
return to the example of Bull Frogs that I used in the lecture on Natural and Sexual Selection. Howard
(1979) demonstrated that male bull frogs tend to mate with the largest male
bull frogs and their is a positive relationship between male bull frog size
and the number of females that a male secures as mates.
Males might choose females on the basis of the quality
of the male or the quality of the territory that the male secures. Let us
return to the example of Bull Frogs that I used in the lecture on Natural and Sexual Selection. Howard
(1979) demonstrated that male bull frogs tend to mate with the largest male
bull frogs and their is a positive relationship between male bull frog size
and the number of females that a male secures as mates.
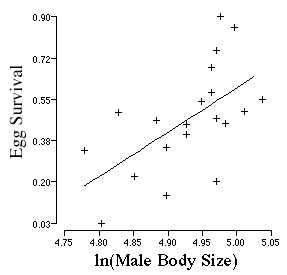 Howard assessed the quality of the male and the male's
territory, by tracking the survival of the eggs through to hatching as a
tadpole. Females have a very strong perference for males that are large
and it also appears that these males are better able to secure a quality
territory with either superior thermal resources which allow for proper
incubation of eggs, or the large males are better able to defend their own
territory against predators and conspecifics that might eat the eggs. Males
of many species participate in such active care of the young and we will
treat this in greater detail in the next lecture. Such aspects of territory
quality can be considere part of the male's extended phenotype.
Howard assessed the quality of the male and the male's
territory, by tracking the survival of the eggs through to hatching as a
tadpole. Females have a very strong perference for males that are large
and it also appears that these males are better able to secure a quality
territory with either superior thermal resources which allow for proper
incubation of eggs, or the large males are better able to defend their own
territory against predators and conspecifics that might eat the eggs. Males
of many species participate in such active care of the young and we will
treat this in greater detail in the next lecture. Such aspects of territory
quality can be considere part of the male's extended phenotype.
Given that the limiting sex can be choosy and the limiting sex is usually the female, I will restrict my discussion of sexual selection to models of female choice. Keep in mind that Roseqvist demonstrated that males choose and models of female choice could be applied to males as well.
Ronald Fisher is generally credited with coming up with model of runaway process in which genes for female preference get tightly associated with genes for the male trait, leading to a runaway that favors ever more elaborate male traits despite their dibilitating effects on male survival. In this sense female choice can lead to maladaptive evolution. Even in the early evolution of the trait, the runaway sexual selection can be so strong that it favors male traits which can be deleterious.
Ronald Fisher also came up with the alternative explanation for the evolution of female choice that has to do with the male trait being an indicator of the male's quality. Such theories are referred to as "good genes" or "indicator models" for the evolution of female choice.
Experiments with female choice seem pretty obvious, however, the contentious nature of the controversy between good genes, and runaway selection makes manipulative experiments critical for determining cause and effect. A female can choose a male on the basis of a number of potential traits, and such male traits might indeed be correlated with one another or indeed, correlated with the female preference per se. It is essential to perform manipulative experiments of the male trait to isolate the effect of a specific male trait on the preferences of females.
Time and again researchers have demonstrated this fundamental observation in female choice experiments. The key is to perform an experiment in which the male ornament is made more elaborate by cutting and pasting a larger ornament on some males and reducing ornament size in other males. A classic demonstration of female choice for supernormal stimuli was carried out by Malte Andersson (1982) in the widowbird. Andersson gave female widow birds the choice of such "cut-em" and "paste-em" males and the females overwhelmingly choose males with the longer ornament.
Many organisms choose mates based on the call, and not the visual system. In such organisms, the ability to use recorded male songs in a "playback" style experiment is ideal because one can remove all aspects of the male phenotype save the salient aspects of the song itself. You can even engineer the song quality and manipulate the same male's song and see how the female choice is affected by such "digitally engineered songs". For example, female crickets prefer a long song type over a short song. The male crickets produce a trill-like song using their legs rubbing against their dome-shaped wings. The dome-shaped wings resonate the song to listening females. A female will tend to walk towards a speaker that is playing a long song compared to an adjacent speaker that is playing a short song.
Females will even choose males on the basis of his extended phenotype. Shrikes are a carnivorous bird of the desert that have the curious habit of skewering lizards onto the thorns of bushes. For years, naturalists used to think that such lizards were used as a food stash. The problem was that the shrikes left the lizards on the thorns until they had crisped up to a tough piece of scales. Now unless skrikes like lizard jerky, something was amiss with the "food larder" hypothesis.
The answer was solved when Yosef manipulated the number of skewered lizards on a male territory. The males with more lizards had females arrive on their breeding territory ealier, and they also got more females than the males with reduced lizard skewers. Presumably females gauge a male or his territory quality by the number of lizards that he has skewerd and such punk lizard jewelry has strong effects on female preference.
Now lets look at the two models descrbing the evolution of female choice, and I begin with a brief review of the finer points of runaway ...
The simplest model for sexual selection, female choice and a male trait assumes a haploid organism (one set of chromosomes). Let us assume that females come in two types and they are genetically based. The male trait is also genetically based:
 Next Generation:
Next Generation:
Notice that the ornamented male has a fitness of 3/2 and the plain male has a fitness of 1/2. So the ornamented male is favored by sexual selection.
The next step in runaway is to consider what happens to both female choice genes and the male trait in their sons and daughters.
Notice that all of the choosy female genes get bundled up with the elaborate male trait.
Lets plot out a table to contrast such associations where:
Choosy females (C) choose Elaborate males (E) and reject normal males (N), and
Non-choosy females (+) choose E and N in a 50:50 ratio,
with what would be expected under random mating (e.g., females come in A and + types but they mate randomly):
Male Matings and Choosy Females |
Male Matings Expected by Random Female Choice | ||||||||||||||||||||||||||
|
| ||||||||||||||||||||||||||
Now the key step is to predict the gametes types in the next generation:
The sons and daughters have genes for the male trait, and both sons and daughters also carry a preponderance of the choosy gene (note that based on the probability of choice, a male receives 1 share of his matings from choosy females, and 1/2 share of his matings from non-choosy females. Thus, from the point of view of the male genes and progeny, 66% of the male Elaborate trait genes in the population are associated with the female trait gene. However, only 33% of the Elaborate trait genes are associated with the non-choosy female gene.
Note also that all drab males breed with non-choosy females and thus 100% of the the drab genes (all of them) are associated with non-choosy females (conversely 0% of the drab genes are associated with choosy females.
Because the combination of Elaborate/Choosy is in excess, more offspring are produced with this combination. This occurs every generation and the choosy females and Elaborate males both benefit and enjoy high fitness.
Both of these facts mean that choosy genes and elaborate male genes occur in a higher frequency than one would expect if the population were mating non-randomly. The central prediction of runaway process is that female choice and the male trait should become genetically correlated or the choosy gene becomes genetically linked to the non-choosy gene. It is more likely that you find a choosy gene associated with the male trait, or vice versa that you will find a non-choosy gene associated with the drab male gene.
What happens each generation is that both the gene for female choice and the male trait become correlated. Note that 1/2 the progeny have both genes for choice and the exaggerated trait.
If we continue this process, one more generation, all of the daughters of the choosy females have both the choice gene and the male trait gene. This means that females will be producing sons and daughters with both choice and exaggerated trait genes together.
Because the exaggerated males have an advantage in subsequent generations and because the choosy female genes are genetically correlated with the male trait, Female Choice and the male trait spread, and they spread together, almost like a wild fire, or as Fisher termed a Runaway Process.
More importantly the genes for female choice and the genes for the male trait become linked in the progeny (even though they are not on the same chromosome). Such linkage by assortative mating leads to the runaway, because the better males who are chosen with high frequency produce lots of daughters who are likewise quite choosy.
A critical prediction of Runaway process is that female choice should be genetically correlated with the genes for male choice.
Tests of this first simple prediction have taken two approaches. First search for the proper pattern between individuals expressing female choice and the male trait. Because, no individual can simultaneously express female choice and the male trait (hermaphrodites need only apply), the test for a genetic correlation between genes for choice and the male trait looks at the correlation between sibs -- daughter's choice and son's trait. Another approach would be to perform and artificial selection experiment in which the researcher selects on females with a given choice and observes the effect of such a choice on the correlation between female choice and the male trait that was not under direct artificial selection.
<picture>Bakker (1994) measured the preferences of Daughters and Sons Coloration and found such traits to be strongly correlated. The daughters and sons tend to receive genes for preference and the male trait from their mothers or fathers. The genes that they receive are correlated from previous generations of selection. This is very strong comparative evidence that traits under sexual selection in stickleback are correlated genetically. This pattern is consistent with the hypothesis of runaway sexual selection.
Anne Houde (1994) found similar effects in Guppy's which likewise have elaborate coloration in males (e.g., reds, blues, black spots), and different kinds of female preference in the female. Anne began with populations of female guppies that had a variety of preferences, and selected males to breed with the females based on the color of their bodies. She controlled sexual selection artificially by allowing certain color types of males to be more successful at mating than other color types. Because the male trait was not under direct artificial selection, the correlated evolution of these traits arises because the female choice genes get bound to the male trait genes by the process of correlated selection. This is very strong experimental evidence that the genes for female choice get associated with genes for the male trait by the process of Sexual Selection.
What happens to female preference in populations of guppies?
There are now a vast number of good genes models on the market. The feature that they all share in common is the notion that the female chooses males on the basis of elaborate ornaments because such ornaments are strong indicators of the superiority of the male's genotype for genes unrelated to the trait. Presumably, a male with an elaborate male trait would have to be quite superior to be able to survive to reproductive age with the large trait and thus he should have many genes that are related to his overall vigor, that are unrelated to ornament per se. Variation in the indicator trait does not have to be genetic, it just should indicate the males physiological vigor or other measure of male quality.
A female that chooses such a male endowed with lots of good genes will also pass on these good genes to her progeny. Traits that aid in the survival of a male will undoubtedly be valuable resource to pass on to her progeny, thus her choice is purely adaptive in that it leads to strong material benefit to her own offspring.
Another term that has been applied to good genes models is the handicap principle. Zahavi proposed the only males in superb condition could carry the handicap that a large expensive ornament would entail.
Marion Petrie (1994) set out to test the idea that females might choose males on the basis of some sexually-selected trait, and her choice of ornament might have an indirect genetic benefits for her offspring.
Again the problems of correlation versus causation come into play. Might a female who has a tendency to search out the best male (not an easy task), is also a robust specimen. She might also have above average genes compared to other females.
Petrie surmounted this difficulty by first establishing which males the peahens like to choose. Based on such choice experiments, she identified the chosen males prior to performing a controlled breeding. She did not let the females choose their mates, rather, she arranged the matings randomly. But she already new that females overwhelmingly preferred the males with the largests spots on his tail.
Petrie found that both of her predictions were borne out.
Male fathers with the largest spots on their tail ornament tended to produce chicks that had both high growth rate and high survival during their residence in Whipsnade Park.
Petrie, M. 1994. Improved growth and survival of offspring of peacocks with more elaborate trains. Nature 371:598-599
Hamilton and Zuk proposed that species under strong sexual selection should be more likely to be parasitized compared to non-showy species. Why should this be so?
They reasoned that males in a species under sexual selection would be selected for the male trait to such an extent that it would begin debilitating their physiological state, which would make them more susceptible to parasites. Not all males in the species would be so affected, and those that had the brightest ornaments would presumably be those males which were also parasite free, and had genes that might allow for resistance to parasitism. Females that chose to mate with the brightly ornamented males would also be acquiring "parasite resistence genes" for their offspring.
Hamilton and Zuk tested their ideas by performing a comparative experiment in which the ranked a male's of each species for showiness from 1-6 and correlated "showiness" (e.g., degree of sexual selection) with data on the risk of being parasitized in each species. The found a very strong correlation between parasites and showiness. However, other studies have failed to find a correlation.
Why are results from their study difficult to interpret?
Well, showy species also tend to be the species in which males are highly polygynous (many female partners per male). Non-showy species tend to be monogamous (usually one female partner/male). If the risk of infection by sexually transmitted diseases is a function of the mating system, then one would expect polygynous, showy species to more likely to have sexually transmitted diseases compared to the more mate faithful monogamous species.
Anders Moller improved on correlational aspects of the above comparative study by performing a careful series of manipulative experiments. Moller reasoned that if the Hamilton-Zuk Hypothesis were true, then the following patterns should be seen within a single species:
In an elegant series of experiments on barn swallows, Moller found support for all four predictions.
1) He applied blood-sucking mites to the offspring in some nests, and these offspring fledged at a smaller size. Fledging size is directly related to survival to maturity.
2) Males vary in the number of parasites at the beginning of the season. To establish that such variation is heritably transmitted to offspring, Moller carried out a cross fostering experiment. He split the clutch that a father sired and placed it in the care of "foster parents". He then innoculated offspring in foster nests with mites to see if they could resist the mites. This tests for the effect of rearing environment on the susceptibility to mite infestation in offspring (e.g., foster nest), compared to the genes that the father passed on for parasite resistance. It covercomes the problems of a father passing on an infestation to offspring.
Offspring that overcame the mite infestation were more likely to come from males that were parasite free at the beginning of the season compared to offspring that remained infested that came from infested fathers.
3) Infestation during youth has an effect on ornament at maturity. By killing mites on some offspring using Pyrethrin, Moller was able to produce males that had longer tails at maturity.
4) Finally, females prefer males with long tails the indicator of a parasite-free male. In cut-em and paste-em experiments, females overwhelmingly prefered males with experimentally elongated tails.
In recent years researchers have been searching for more and more attributes that might indicate mate quality to a choosy female. One simple trait that has been used is the idea of fluctuating asymmetry in the ornaments that males present to females. Fluctuating asymmetry refers to random perturbations in ornament size on one side of the body axis but not the other. Biologists hypothesize that such random perturbations reflect some kind of developmental instability.
Researchers believe that asymmetry indicates genes that aren't working well with one another -- bad epistatic interactions. Thus, a female that mates with a male that is assymetric in in their ornament may be choosing a male that has genes that are not working well together, compared to a female that chooses a very symmetrical male.
Let us consider the molly. Male mollies not only have a sword on their tail that is a sexually selected trait, but they also have strong vertical barring on their sides. My colleague Molly Morris did a female choice study on swordtails and she was interested in whether female mollies choose males on the basis of the number of bars on one side of the body versus the other. To perform the choice experiments, she used a small piece of dry ice to make males assymetrical- almost like removing a tatoo in a tatoo parlor. Females overwhelmingly choose males that had symmetric bars on one side of the body versus the other.
This is a relatively new idea and it is very similar to the idea of unconcious selection. Before discussing the idea of indirect female choice let us think of a simple model of active female choice.
It is very difficult to shy away from the idea that females (or males) are somehow making active cognitive choices regarding the mate quality. Recall the three criteria that I had for cognitive thought processes in animal foraging (see the bumble bee lectures).
Cognition is defined in terms of three items (Real 1993):
Clearly females are surveying males and they are taking in information. But showing that such information is stored in memory.
E.g.,
Ornament 1
Ornament 2
Ornament 3
....
Who should I choose, hmmm?? Given a choice of "n males" a female should choose the best of n that she has encountered.
This kind of problem has yet to be answered in full detail. Clearly females are making such cognitive choices, showing how it works remains unanswered.
Another idea of female choice and sexual selection has just surfaced and it is called indirect choice. It is the notion that females are not making active choices of males, but through their actions, they are selecting for sexually selected traits. Many courtship routines entail elaborate and vigorous chasing and the like, and a female may indirectly select the best male by performing such actions. Only the fitest male can keep up with her and she will weed out any stragglers. Selection favors the more vigorous males and any aspects of secondary sexual traits that aid in such pursuit.