Back to the Syllabus
Behavior and Speciation Mechanisms
Barry Sinervo©1997
Index
Biological Isolating Mechanisms
Modes of Speciation Classified by Geography
Clinal Variation
Allopatric Speciation
Parapatric Speciation and Allo-Parapatric Speciation
Ring Species in Gulls
Paint Experiment in Gull Species Recognition
Ring Species in Salamanders and the Evolution
of Mimicry
Sympatric Speciation
Polyploidy
Reproductive Character Displacement and Mate
Calls in Frogs
Production of Asexual Parthenogenetic Species
Multiple-Niche Polymorphism in Insects and Host-Plant
Preference
Allopatric versus Sympatric Speciation: Case
Studies
Conditions for Sympatric Speciation -- Assortative
mating
Cultural Inheritance of Song and Speciation
in Darwin's Finches
Deme Recognition and Female Mate Choice
Founder Effect Speciation
Runaway Process, Sexual Selection, and Speciation
Species are groups of actually or potentially interbreeding populations,
which are reproductively isolated from other such groups.
Mayr 1942
Biological Isolating Mechanisms
Ernst Mayr's formulation of the biological species concept clarified
a vexing question for evolutionary biologists: What is a species? Mayr's
view of the species concept has widespread acceptance owing to its simplicity.
However, how species arise by the process of natural selection remains a
source of great controversy. The problem is that the process of speciation
takes a very long time to complete, and it occurs on a very large geographic
scale. In addition to Mayr's definition additional concepts are quite useful
in delimiting species.
Hybrid sterility definitely separates two species, although it
is not the sole criterion. Hybrid sterility is a special instance of reproductive
isolation. In the cases of Reproductive isolation between species the
barriers to mating do not just have to be due to incompatibility between
genomes. The blocks could be due to behavior or morphology. These isolating
mechanisms which serve to limit gene flow between species are categorized
in terms of pre- or post-mating mechanisms:
A) Premating isolating mechanisms -- prevent union of gametes
-> zygote
- mates do not meet (seasonal or habitat isolation)
- mates meet but do not mate (ethological or behavioral isolation)
- mates meet but no sperm transfer (mechanical isolation)
B) Post-mating isolating mechanisms -- varying degrees of hybrid
sterility or fitness
- sperm transfered but dies before fertilization
- zygote dies
- zygote produces an F1 adult that has reduced viability (survival)
- hybrid is viable, but partially or completely sterile (fecundity) or
the F2 is deficient.
The answer to the adaptive value of pre- versus post-mating isolating
mechanisms is revealed by asking the following questions. Which of the two
mating isolation mechanisms is the most effective in reducing the risk of
producing low fitness offspring? Which mechanisms is most efficient in terms
of time and lost reproductive opportunity?
An organism that mates with a different "semispecies" may produce
low viability offspring and this individual would have lower fitness
compared to an organism that discriminates against such semispecies and
mates with members of its own semispecies. The evolution of species discrimination
mechanisms are clearly an advantage over one that only possess post-mating
isolating mechanisms. Such behavioral descrimination ability should evolve
rapidly in the area of incipient speciation, under the right conditions.
Biological barriers to interbreeding undoubtedly arise during the process
of speciation because such blocks have selective value. Behavioral isolation
can be the strongest form of reproductive isolation. The clearest examples
of biological species concept in action are provided when two species which
have overlapping geographic ranges do not interbreed owing to differences
in pre-mating behaviors. One species or both species refuse to engage in
the mating rituals because the stereotyped mating rituals are to different.
Species might simultaneously possess both premating and post-mating isolating
mechanisms in which case the probability of interbreeding is even more remote.
What are the right conditions? This is a very difficult question to answer.
It requires an understanding of the field of population genetics and the
crucial parameter is how much migration occurs from areas where each
semispecies exists in its pure form. A second parameter, has to do with
the degree of genetic similarity between the two pure semispecies.
Migration is the movement of animals between what are referred
to as populations of interbreeding animals. Rates of migration are usually
expressed as the proportion of individuals in a population that disperse
from one population to the next each generation. If migration between the
region of contact between the two semispecies and the rest of the semispecies
ranges is too high, then the two semispecies will become homogenized into
one species even if such interbreeding results in lower fitness because
of hybrid unfitness. If migration is low enough, then the two pure semispecies
will eventually become differentiated into two species owing to the evolution
of isolating mechanisms. Evolution of such isolating mechanisms is contigent
upon the degree of genetic similarity between the two semispecies.
 Large differences in genetic similarity between
semispecies are more likely to produce unfit hybrids than small differences
in similarity. If the two semispecies have many genetic loci that differ
in the kind of allele that they possess, then those alleles might not work
very well together, and more importantly the way two different genetic loci
work together may begin to breakdown. This is referred to as genome-wide
genetic interactions. The correct operation of one genetic locus (lets
say an enzyme) may require a particular form of the enzyme that is produced
by a different locus (e.g., epistasis).
These enzymes are most likely to be compatible in their function if they
come from the same species. If they are from different species, they may
not integrate well with other loci in the organism and the hybrid individual
may die. Imagine such genetic interactions occuring at all possible loci
(e.g., 10,000 loci in Drosophila or perhaps 100,000 loci in humans). The
probability of a hybrid being more unfit has to do with how many loci in
each semispecies are homozygous for different alleles. Hybrid breakdown
occurs when many of these loci are fixed for different alleles in the two
subspecies and the production of offspring by the hybrids is not possible
because and F1 X F1 produces too many incompatible gene by gene combinations.
Large differences in genetic similarity between
semispecies are more likely to produce unfit hybrids than small differences
in similarity. If the two semispecies have many genetic loci that differ
in the kind of allele that they possess, then those alleles might not work
very well together, and more importantly the way two different genetic loci
work together may begin to breakdown. This is referred to as genome-wide
genetic interactions. The correct operation of one genetic locus (lets
say an enzyme) may require a particular form of the enzyme that is produced
by a different locus (e.g., epistasis).
These enzymes are most likely to be compatible in their function if they
come from the same species. If they are from different species, they may
not integrate well with other loci in the organism and the hybrid individual
may die. Imagine such genetic interactions occuring at all possible loci
(e.g., 10,000 loci in Drosophila or perhaps 100,000 loci in humans). The
probability of a hybrid being more unfit has to do with how many loci in
each semispecies are homozygous for different alleles. Hybrid breakdown
occurs when many of these loci are fixed for different alleles in the two
subspecies and the production of offspring by the hybrids is not possible
because and F1 X F1 produces too many incompatible gene by gene combinations.
Genetic similarity drops as we change from the different levels
of differentiation, going up the heirarchy of differentiation: populations
-> races -> subspecies -> semispecies -> sibling species ->
species. By the time species are compared there is generally little or no
gene flow between species. As genetic similarity drops, the differences
increase and the possibility of hybrid breakdown increases. A species is
thought to have a coadapted genome in which many genes are finely
tuned by the proper interactions with other genes.
Clearly genetic differentiation is occuring during speciation but it
is not enough to define a species. It can vary from organism to organism
and from mode of speciation to the next. In practice, even the criterion
of no gene flow between species is not the defining feature of species.
As we will see below, the biological species concept begins to break down
in areas or at times of speciation or incipient speciation (semispecies).
In such cases, alleles can leak across the hybrid zone from one species
into another.
Index
Modes of Speciation Classified by Geography
To understand the origin of species differences it is necessary to consider
how genetic differences arise between species. Geographic subdivision decreases
gene flow. Depending on the population size, genetic drift can play a major
role in promoting genetic differentiation. If population size remains low
for a long period of time it is possible for genetic differences to accumulate
between geographic areas by the random process of genetic drift. Selection
can promote speciation because genetic changes can rapidly accumulate in
one area where selection favors certain behaviors and morphology relative
to another area where alternative alleles are favored by natural selection.
Finally, the amount of gene flow which is usually described in terms of
the migration rate is a key parameter that influences the rapidity of speciation.
By lowering gene flow, geographic subdivision promotes the process of speciation.
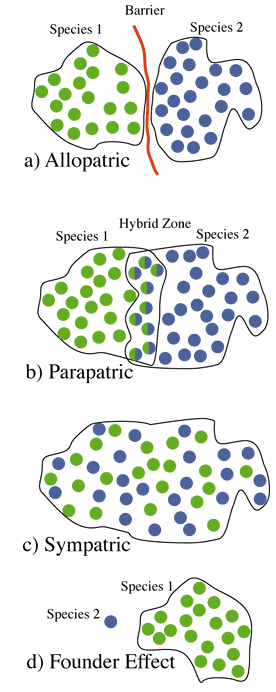
Models of speciation relate to the degree of geographical subdivision
for Allopatric -->Allo-parapatric --> Parapatric --> Sympatric
which ranges extreme to none. Another parameter that can be used to descriminate
the types of speciation is related to rates of gene flow and how alleles
at many genetic loci are distributed across the species range.
- Allopatric Speciation -- speciation occurs in geographic isolation,
- Founder Effect Speciation -- a special kind of allopatric speciation
in a small isolated population on the edge of a species range
- Parapatric Speciation -- speciation in adjacent populations
with gene flow,
- Allo-Parapatric Speciation -- populations initially separated
(allo-) but then secondarily come into contact with subsequent parapatric
speciation,
- Sympatric Speciation -- speciation within a panmictic population.
Figure 2. Models of speciation based on geographical subdivision.
Index
Diagnosing Models of Speciation from Clinal
Variation
Patterns of variation across the species may provide some basic diagnostic
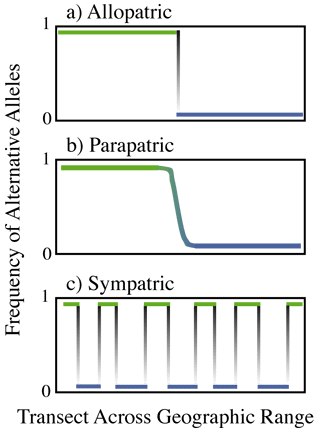
features that distinguish the four modes of speciation. A cline is any systematic
change in a trait across the range of a species. Clines can be abrupt or
gradual. Presence of clines in morphological or behavioral traits are used
as evidence for one mode of speciation over another (see Figure 3).
- Presence of a cline would suggest a particular mode of speciation
has operated in the past. Presence of a geographical barrier at the cline
would reinforce the notion that the speciation event was due to allopatric
speciation.The change in behavior traits, morphological traits, allozymes,
or some other form of genetically-based variation resembles a step function.
A very abrupt change in traits where the two species come into contact
would suggest little or no gene flow in the present day. Lack of gene flow
in the present day could be could be due to the evolution of reproductive
isolation mechanisms during allopatric speciation in the remote
past and the original barrier has been removed.
- The Shape of the Cline might also be indicative of gene flow.
Clines can be gradual across a hybrid zone and span 1000 of km or clines
can be very abrupt and occur over a few 100 m. Is the cline maintained
by persistent selection that favors one form of the species on one side
of the cline and another form on the other side of the cline? If yes, then
the two species along with their hybrid zone might be a candidate for the
parapatric model of speciation. In the case of a cline that is being
maintained by persistent selection, the cline may have arose as a primary
response to different selection regimes in the species range.
- Origin of the cline or hybrid zone is another attribute that
might distinguish models of speciation. Is the cline due to secondary contact?
In this case, the two species were in allopatry in the remote past and
brought into secondary contact and weak gene flow began once the sibling
species were rejoined at the hybrid zone. However, complete reproductive
isolation did not occur during allopatry, but rather the the species differences
were reinforced once the two species came into secondary contact. The cline
is said to have evolved by allo-parapatric speciation.
- If there is no cline and the two species can be found co-occuring
across a large geographic range, sympatric speciation may have produced
behavioral isolation. A transect across the range of the two species would
show isolated pockets of one pure genotype, or the other pure genotype,
with no present day gene flow between the two species.
Figure 3. Hypothetical change in frequency of allele frequency
across the range of two species under allopatric, parapatric, and sympatric
models of speciation.
Index
Allopatric and Parapatric Speciation
Any geographic factor that severely restricts gene flow will allow populations
of a species on one side of the barrier to begin to differentiate from species
on the other side of the range. Given enough time, the genetic differences
became large enough to warrant calling the "races" on one side
or the other a different species.Allopatric speciation occurs under the
following conditions:
- isolation of a colony (e.g., island and mainland)
- division of range by extrinsic barrier (e.g., mountain range) or extinction
of populations in the intermediate parts of the range.
- isolation by geographical distance.
Parapatric versus Allo-Parapatric Speciation
In order for a species to arise by parapatric speciation, there
must usually be a very strong change in the environment. Natural selection
on one side of the environmental transition favors one set of traits compared
to the traits favored on the other side. The existence of clines as evidence
for parapatric speciation is generally not considered very good evidence.
However, if selection is strong enough it leads to disruptive selection
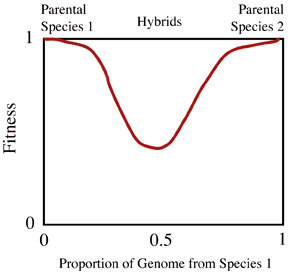
at the abrupt transition that parapatric speciation occurs. The fitness
of an individual is determined by the fraction of genes from each parental
type. Individuals that are pure for each parental type will tend to have
higher fitness than individuals with a genome that is formed from a mixture
of the two parental combinations. These hybrids come from crosses between
the two parental forms. However, animals that interbreed (e.g., mate with
a different type) at such hybrid zones tend to show underdominance
in fitness, that is their progeny the heterozygotes (hybrids) have lower
fitness. Another way to think of this pattern is to view it as disruptive
selection against individuals heterozygous for a large number of loci. The
genes in the heterozygous condition have lower fitness than homozygous configurations
because of the strong environmental differences on either side of the contact
zone.
If there is any viability of the hybrids, alleles may leak across and
introgress across the zone. Leakage of alleles across the hybrid zone is
referred to as Introgressive hybridization. Some alleles may spread
quite far across the zone whereas others will be quite abrupt and coincide
with the hybrid zone.
The big question, can we distinguish between primary and secondary contact?
In principle, we cannot ,which is the case and that is why the models of
speciation are so fiercely debated. The process of speciation is by its
very nature a historical process. The origin of the cline is a key component
of all all models of speciation, and this event can only be inferred from
the patterns of clinal variation that we see across the present-day species
ranges. We see many fully formed species, and while we also see incipient
species in the process of formation, the actual cases are difficult to resolve
as allopatric versus parapatric speciation, because events have occured
in the remote past.
Nevertheless, studies of the geographic pattern of races in a species
and their likelihood of interbreeding have provided us with splendid examples
of apparent allopatric speciation. Nothing is more challenging to the imagination
that a special process of allopatric speciation which produces a ring species.
Index
Ring Species in Gulls
 Ring Species arise from a peculiar form of allopatric
speciation that takes place when the center of a species range is unoccupied
because the habitat is unsuitable. Adjacent races all around the ring will
interbreed, however the races at the termini are so divergent, that there
is no interbreeding, and they can exist in sympatry without interbreeding.
Ring Species arise from a peculiar form of allopatric
speciation that takes place when the center of a species range is unoccupied
because the habitat is unsuitable. Adjacent races all around the ring will
interbreed, however the races at the termini are so divergent, that there
is no interbreeding, and they can exist in sympatry without interbreeding.
The species range of Sea Gulls in the Arctic Ocean represents a classic
example of a ring species. The polar ice cap limits the species range of
Sea Gulls to a circumpolar ring.
There are a number of races of Sea Gulls as one traverses around the
globe. Races from America freely interbreed with races from Europe. Races
from Siberia freely interbreed with races from America. Races from Siberia
freely interbreed with races from the Caucauses. However Central European
races do not interbreed with those from Western Europe. All along the ring
there is gene flow, but where the two ends of the ring meet in Europe there
is no gene flow. The ring species of gulls forms a cline in that there are
small changes in coloration and behavior of the gulls along the length of
the ring. However, not enough mate discrimination occurs between adjacent
populations until the ends of the ring meet in Europe.
A Paint Experiment in Gull Species Recognition
Work by Smith on a different species of sea gull demonstrates that the
behavioral isolating mechanism which operate in gull species recognition
may be quite simple. Smith investigated the signal that is used to isolate
the two species into Larus hyperboreus, the glaucous gull and Larus
thayeri, Thayer's Gull. The glaucous gull has a yellowish eye ring,
and Thayer's Gull has a purple eye ring. Smith painted the eye rings of
Thayer's males yellow, and this allowed Thayer males to successful pair
with glaucous females. However, the males would not mate with the glaucous
females until Smith painted the glaucous females purple. Such paint experiments
are a powerful means of demonstrating the meaning behind signals and the
role in species recognition.
Index
Ring Species in Salamanders and the Evolution
of Mimicry
A ring species of salamander, Ensatina scholtzi, is found in higher
altitude regions of California such as the sierra and coast ranges, but
is conspicuously absent from the great Central Valley of California. Abundant
evidence of gene flow along the cline is provided by comparing the frequency
of electrophoretic alleles in adjacent populations of the salamander. Gene
flow is present throughout the coast range from south to north, from the
coast range to the ranges that cross northern California to the sierras,
all down the length of the sierras, from the sierras to the transverse ranges
of california, but gene flow abruptly ends in a sharp dividing line located
in southern California. The ring of gene flow that traverses thousands of
miles around the perimeter of California finally expires and speciation
appears to have run its course.
The ring species of salamander is also noteworthy in terms of its morphology
and behavior. Where Ensatina co-occurs with the California Newt in the coast
range, Ensatina has evolved a color pattern that resembles the toxic Newt
in several regards. It has a reddish cast to its belly, and its eye is colored
a brilliant yellow, both are features that are thought to be warning coloration
in the newt. In Ensatina, the coloration appears to be a ruse as Ensatina
is not at all toxic. We will take up the causes of such warning coloration
in chapter 12 which deals with predator and prey
interactions.
Index
Sympatric speciation
Sympatric speciation is the origin of two new species with the two forms
in the same place. Conditions for speciation by sympatric speciation are
very stringent. Gene flow between incipient species must be eliminated or
severely curtailed, or reproductive isolation is not possible. Whereas isolation
by geographic distance as is seen in allopatric and parapatric speciation
can limit gene flow, arriving at sympatric speciation when individuals are
found in the same geographic locale is a challenge. Controversy surrounds
all theories of sympatric speciation that involve a gradual speciation event
in sibling species that are in a panmictic, or can freely interbreeding
across the range.
A key aspect of the theory of sympatric speciation involves the process
of natural selection that favors the evolution of discrimination on the
part of the female and also the divergence of male traits that females use
for mate discrimination. Dobzhansky suggested that natural selection should
favor the more finely tuned discrimination mechanisms of the female, in
response to the reduced viability that results from hybridization. Thus,
a key component of this theory is some form of disruptive selection that
selects against hybrid phenotypes, which then favors evolution of mate discrimination
mechanisms. A process referred to as reproductive character displacement
should accentuate differences in courtship behavior or mate preference in
sympatric populations of the two incipient species compared to the differences
between allopatric populations of the two incipient species. Butlin (1987)
coined the term reinforcement to describe such evolutionary interactions
between incipient species, in which hybrid unfitness promotes divergence
in courtship behavior or mate preference in sympatric compared to allopatric
populations.
The evolution of such divergence, in the face of gene flow is a tremendous
challenge for the evolution of species by the process of sympatric speciation.
Felsenstein showed that the process of sympatric speciation is a fairly
unlikely event. He modeled this process in terms the evolution of two species
attributes: 1) the phenotypic attributes that give rise to hybrid unfitness,
and 2) the genes that give rise to mate discrimination. Generally, the genes
for fitness, are not expected to be linked (either by pleiotropy or by physical
linkage) to the genes that give rise to mate discrimination. This key fact
places a tremendous genetic constraint on speciation by the process of reinforcement
(see Side Box 5.4).
Polyploidy
Despite the apparent difficulties most theories of sympatric speciation,
one form of sympatric speciation is universally accepted -- the instantaneous
mode in which changes in ploidy lead to instantaneous reproductive isolation.
While extremely common in plants, it is less common in animals and well
documented cases are found in the lower vertebrate classes: fish, amphibians,
and reptiles.
For example, Gerhardt has identified a case in which a single species
of Hyla tree frogs has undergone tetraploid formations. The formation
of tetraploid (4n) from the fusion of two diploid parents (2n) leads to
a new species of treefrog that can only breed with another individual that
has similar ploidy (Gerhardt 198X). The fusion of gametes from a diploid
parent and tetraploid parent produces triploid offspring which die during
larval stages and the few hybrids that reach adult size are sterile (Johnson.
While production of triploids can be achieved in the laboratory, triploids
are rarely found in natural populations of Hyla tree frogs (Gerhardt
1982).
Because the diploid parental species achieves reproductive isolation
from the tetraploid daughter species in one fell-swoop, sympatric speciation
is easy to achieve. Indeed Gerhardt and his colleagues have found that the
production of new tetraploid species has occurred several times in this
species complex. Diploid populations of Hyla chrysoscelis probably
gave rise to tetraploid populations of H. versicolor (Tymowska 1991).
Character Displacement in Mate Calling and
Female Preference
Gerhardt has tested female preferences for male call frequencies of the
two species of Hyla frogs in areas of sympatry and allopatry. Males
of each species from areas of sympatry are more divergent in key aspects
of their calls compared to males from areas of allopatry. In areas of sympatry
Hyla chrysoscelis males produce a longer call than co-occuring H.
versicolor. In areas of allopatry the songs of both species are of medium
duration.
The key test of character displacement via reinforcement involves assessing
whether females have evolved more fine scale preferences. The hypothesis
of character displacement would suggest that females from sympatric populations
showed stronger preferences for acoustic stimuli of the males call than
females from more remote populations. Gerhardt carried out a number of phonotaxis
experiments to test whether females in allopatry have evolved more fine
scale discrimination of the species differences in mate call. Gerhardt created
synthetic calls which allowed him to vary specific attributes such as call
length rather than the fine scale aspects of the song. He then played songs
from two speakers. One speaker played a male with the short synthetic call
typical of the males of the females area, the other speaker played a male
with longer synthetic calls. Females from areas of sympatry were much more
likely to discriminate the correct conspecific short song that females from
more remote areas of allopatry.
Index
Production of Asexual Parthenogenetic Species
The production of polyploid species in lizards is also quite common.
For example, species of Cnemidophorus or whiptail lizards from the
southwestern Deserts have produced daughter species by polyploidy repeatedly.
Not only have sibling polyploid species been produced, but reproductive
isolation is achieved by another interesting evolutionary event -- many
of the daughter species that are produced are asexuals which means they
are all female and do not require males to produce offspring. The evolution
of an asexual or parthenogenetic species can eliminate gene flow even more
effectively than ploidy changes alone. If sperm is not required, the newly
derived clonal species instantaneously begins an independent existence.
The asexual daughter species of Cnemidophorus no longer needs males to produce,
however, reproduction in females of the asexual species are facilitated
by male-like behavior in a curious fashion. If females participate in pseudocopulation
in which one female mounts another a female in the stereotypical male-female
copulatory position, the female produce more eggs, and the interclutch interval
is reduced .
Interestingly, the block to the formation of hybrids is not necessarily
absolute in some parthenogenetic species. Genes from a facultative asexual
species could be transmitted to a sexual species. Most of the time the asexual
species can produce parthenogentically. For example, asexual species of
flatworms from lakes in Europe have evolved repeatedly. Flatworms are also
unique in that they are simultaneous hermaphrodites, which maintain active
testes and ovaries. While the testes of the parthenogenetic species are
much reduced, they still produce viable sperm. The asexual species of flatworm
can produce haploid sperm that it can transfer back to the sexual species,
which allows for one way gene flow from the facultatively asexual daughter
species, back into the parental sexual species.
Index
Multiple-Niche Polymorphism in Insects and
Host-Plant Preference
Without strong assortative mating of some form sympatric speciation is
an impossibility. Most theories of sympatric speciation other than ploidy
changes or the evolution of a parthenogenetic species are controversial.
Any sympatric speciation theory that involves a gradual change in sympatry
is difficult because gene flow would swamp out the build up genetic differentiation
that forms the hallmark of speciation. There is another notable exception
which involves the evolution of insect preferences for feeding on certain
species plants. It is thought that the following mechanism may be why there
are more insect species of insects on the planet than any other organism.
If so, then this mechanism is truly important from the point of view of
animal behavior and diversity. Part of the reason why it is so prevalent
is because insects can evolve detoxifying specificity towards their host
plants in a specific enzyme -- P450 (the details of which need not concern
us). However, the acquisition of such specificity should lead to rapid acquisition
of host plant preferences -- a behavioral trait.
One class of models maintains that there arise multiple-niche polymorphisms.
Heterozygotes are inferior because extreme specialization in detoxifying
capability is necessary to successful survive and reproduce on different
species of plants. Assortative mating among homozygotes is a key component
of this theory. This minimally requires two loci to evolve simulatneously,
and thus we establish strong linkage disequilibrium for the niche polymorphism
loci and the assortative mating loci. More loci makes it unlikely to occur.
One classic and well accepted form of sympatric speciation involving
animal behavior has to do with host plant choice by insects. Many host plants
of insects produce very nasty chemicals in repsonse to being eaten. The
chemicals are meant to deter the herbivores, thus in order to continue feeding
on the plants, the insects must develop mutations that allow it to detoxify
the chemicals. Because different host species produce very different toxins,
the mutations that allow an insect to detoxify one host-plants arsenal of
toxins do not work with another. Here are the key issues involved in this
mode of speciation.
An insect must evolve the detoxifying mutations, and also evolve
host-plant preferences that allow it to become very choosy about
the kind of plants it is feeding on. The mutations do not have to occur
simultaneously, but the detoxifying mutation probably arises first
and the behavioral preference evolves as a refinement to the detoxifying
phenotye because individuals that match their own detoxifying abilities
with the proper host plant would be strongly favored by natural selection.
In general, two such mutations are required, because it is difficult
to imagine getting a single mutation that has a pleiotropic effect on detoxifying
ability and host-plant preference. The two mutations should also arise on
the same chromosome in close proximity to one another. This causes the behavior
for host-plant specificity to be tightly correlated with the detoxifying
genes. The linkage between the two loci forms a genetic correlation
between the behavioral and physiological traits. An insect that ends up
with the wrong allele for detoxifying matched up with the wrong preference
is in trouble (recall the problem for stripped and spotted snakes). By having
the mutations on the same chromosome, a super gene for the detoxifying
ability and host-plant preference is produced. It is possible for the genome
to become rearranged after the mutations arise. This would require a third
chromosomal rearrangement mutation which makes it less likely.
Technically, the incipient insect species which feeds on a new host plant
species is found in the same area as the ancestral species, but by virtue
of different host plant preferences and mating microhabitats, they no longer
exchange genes. The easiest way for many insects to find suitable hosts
for their own progeny would be to oviposit their eggs on the host plant.
If you oviposit on your host plant, chances are mating would also occur
on the plant prior to oviposition.
This speciation process not only leads to adaptive plant choice, but
because other members of the insects own incipient species also choose the
same plants, the insects will mate with like members of their incipient
species and not with other members, and reproductive isolation is achieved.
Sympatric versus Allopatric Speciation
Assortative Mating is a Stringent Condition
for Sympatric Speciation
The formation of new sexual species by changes in ploidy and the formation
of asexual species are both examples in which the new species becomes reproductively
isolated from the parent. How likely is sympatric speciation in a group
if there is any gene flow between incipient species. Lets re-visit an old
system -- the beak morphs of African seedcrackers (Smith) and speculate
about what might happen to Pyrenestes in the not too distant future.
If S (small-billed) birds are much more successful in feeding on small seeds
and L (large-billed) birds are much more successful in feeding on large
seeds, why shouldn't these two morphs begin to form a new species, each
specializing on a specific species of sedge? Natural selection on the two
morphs appears to promote character divergence. Selection provides a key
route for the rapid accumulation of genetic differences.
However, based on the calculations of observed and expected proportions
of matings between small and large-beaked morphs, we have already seen that
seedcrackers mate randomly (see Side Box 2.4).
In order for there to be even a remote chance of speciation in seed crackers
they would have to mate assortatively. Without assortative mating the gene
flow between the finch morphs would continue unabated.
Table 5.1. Observed frequency of large and small-beaked morphs
is random (A) which contrasts strongly with patterns expected under strong
assortative mating (B)
| A) Observed random mating in seed crackers. |
B) Assortative Mating required for speciation. |
| |
female of the the pair |
S |
L |
| male in the pair |
S |
34 |
14 |
L |
14 |
6 |
|
| |
female of the the pair |
S |
L |
| male in the pair |
S |
48 |
0 |
L |
0 |
20 |
|
Index
Cultural Inheritance of Song and Speciation
in Darwin's Finches
The natural selection observed in seed-cracking African Finches may form
a model for how natural selection has acted on feeding behavior and performance
of Darwin's Finches on the Galapagos Islands. For Darwin, the Galapagos
islands shaped his thinking like no other place on the planet. These islands
continue to be a source of inspiration for field biologists interested in
the evolution of behavior. Peter and Rosemary Grant, Price, Boag and Schluter
have carried out a series of elegant studies that document selection on
different species.
Each of the twelve species of Darwin's Finches have evolved specialized
morphologies and behaviors that are adapted to their ecological niche. On
some islands one finds two or three species with drastically different beak
morphologies that they use in feeding. One species of finch has even evolved
a woodpecker lifestyle in that it feeds on insects under bark. Interestingly
this species does not use its bill to extract the insects, rather it has
evolved tool using behaviors. The species picks up twigs that it uses to
probe the nooks and crannies of bark and extracts insects. The males of
each species also have unique species-specific songs, and females strongly
prefer song types of their own species over other species.
Most of the larger islands in the Galapagos archipelago have several
species of finches which raises questions regarding the speciation events
that have produced the diversity of finch species. Have the species of Galapagos
finches evolved by sympatry on the same island? Alternatively, have the
species evolved by founder effect speciation or speciation in allopatry
on adjacent islands. To get more than one species on each island requires
subsequent invasion events between islands where one species disperses to
another island where a second species is found. Alternatively, The dispersing
species then colonizes successfully and begins to grow in population size.
The two species begin competing on some of the same resources, however,
by a process of character displacement each species evolves specialized
behaviors and morphology that allow the species to specialize on certain
resources such as plants with different sized seeds.
A possible mechanism for the evolution of species in sympatry would be
the origination of a gene of major effect that causes a new beak morph to
evolve. Each morph would feed on a slightly different resource, much like
the morphs of seed-cracker in Africa. However, matings between morphs of
seed crackers from Africa produce perfectly viable young, and these morphs
are considered a single species. In addition there are not hybrids in seed
crackers as heterozygotes are hidden by the dominant effect of the Large
beak allele. Finally the lack of assortative mating between morphs of African
finches suggest that speciation is unlikely. Available data on the success
of hybrids between different species of Galapagos finches suggests that
hybrids have high viability (Grant and Grant 1996). Moreover, the hybrids
are intermediate in terms of many morphological traits, including beak morphology.
One of the key requirements for sympatric speciation would be a lower fitness
of hybrids that would then favor the evolution of discrimination mechanisms
between the two sympatric species of Galapagos finches. Where hybrids are
formed between species, the hybrids appear to quite viable. In the absence
of such a fitness cost, it is unlikely that species recognition mechanisms
evolved in sympatry.
The evolution of the species in allopatry is a much more likely mechanism
in light of the lack of hybrid inviability. On each island, species would
have acquired differences in morphology, but also important differences
in song also arise in allopatry. When the dispersal of birds occurs among
islands, gene flow is minimized because of evolved differences in song type
among males, and because females tend to choose males on the basis of song.
In order to understand, speciation mechanisms in Darwin's Finches we need
to understand the transmission of song between generations.
Figure. X Similarity of song between father to son, and between
paternal and maternal grandfathers to son.
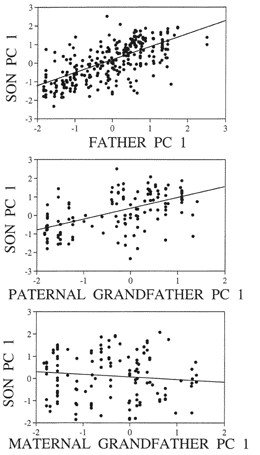 Darwins finches transmit song from father to son. Morevoer,
the transmission of song from father to son is likely to be cultural and
not genetic. If song is genetic, we would expect that a male bird could
transmit genes for song to their daughter. While daughters do not sing their
grandfathers song, they would still be expected to pass the genes for song
on to their own progeny. Consequently, we would expect there to a positive
correlation between maternal grandfather's song and the mother's son, but
there is no relationship. In constrast there is a relatively strong and
positive correlation between the paternal grandfather's song and the son,
and even a stronger correlation between the father and son. This indicates
that song has a patrilineal inheritance, and moreover, is culturally transmitted.
Darwins finches transmit song from father to son. Morevoer,
the transmission of song from father to son is likely to be cultural and
not genetic. If song is genetic, we would expect that a male bird could
transmit genes for song to their daughter. While daughters do not sing their
grandfathers song, they would still be expected to pass the genes for song
on to their own progeny. Consequently, we would expect there to a positive
correlation between maternal grandfather's song and the mother's son, but
there is no relationship. In constrast there is a relatively strong and
positive correlation between the paternal grandfather's song and the son,
and even a stronger correlation between the father and son. This indicates
that song has a patrilineal inheritance, and moreover, is culturally transmitted.
Figure X. Transmission of songs from father to son in Geospiza fortis
across five generations. While most males copy their father's song (right
hand lineage), some copy the song of another male.
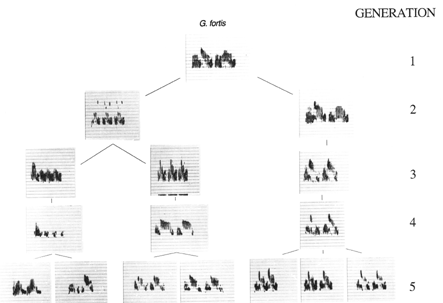 The work of the Grants on the Galapogos Islands demonstrates
that the transmission of dialect occurs in a paternal lineage by the process
of cultural evolution. Mistakes are occaissional made in the transmission
of song from father to song. In such cases the progeny may learn a song
from a neighboring male that is from a different species and such misimprinting
may increase the likelihood of a male of one species pairing with a female
of the other species.
The work of the Grants on the Galapogos Islands demonstrates
that the transmission of dialect occurs in a paternal lineage by the process
of cultural evolution. Mistakes are occaissional made in the transmission
of song from father to song. In such cases the progeny may learn a song
from a neighboring male that is from a different species and such misimprinting
may increase the likelihood of a male of one species pairing with a female
of the other species.
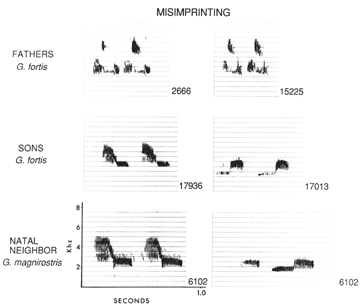
The cultural transmission of song has several important consequences
for speciation mechanisms in Darwin's Finches. First, cultural transmission
of song may allow for misimprinting of song to occur. If sons learn a song
during a critical developmental period, and the father dies prior to this
imprinting period, a nieghbor may fill in the role of song tutor, and the
son learns the neighbors song (see Figure X). In some cases, the juvenile
birds learn the song of another neighbor that is a different species. While
rare, such cases of misimprinting allow for genes to flow from one species
of sympatric Darwin's finch to another. Finally, While females of Darwin's
finch do not learn sing a father's song, they learn to recognize the song
of the father and prefer to mate with males that sing the species specific
song that they learned during their youth. Most of the time, species of
Darwin's finch correctly learn song as males, and females correctly choose
males as mates based on their song. However, in rare cases mistakes are
made, and some hybridization occurs.
Figure X. Misimprinting of song in Geospiza fortis sons that had G. magnirostris
neighbors.
Index
Deme Recognition Female Choice for Natal
Song
An additional requirement for speciation would be selection on females
to strongly prefer males with the song that they heard their fathers sing.
A complete answer to this question requires and experimental approach, which
owing to the sensitive nature of Galapogos Islands, is impossible to perform
on Darwin's Finches. However, many birds are only sexually responsive to
mates from their own natal areas. The following experiment illustrates
that female birds learn songs of appropriate mates in their youth and then
choose rather males that
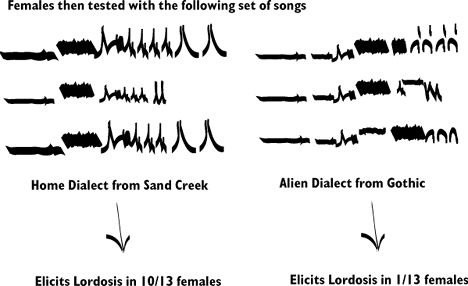
Baker first used tudor males to teach young birds a song dialect. Baker,
found that female white-crowned sparrows responded positively to male courtship
songs at a higher rate if the song was derived from their home dialect compared
to an alien dialect.
The key feature of this study demonstartes that female birds learned
the song during the nestling phase. In addition, it is possible to fool
females into preferring a truly "non-natal" song, or one from
a region other than their true source population. By playing them a song
that is from a different source population during the critical song learning
period. For example, if tudor songs had been from Gothic the females should
have preferred Gothic males at maturity and rejected Sand Creek Songs.

Songs similar to the tudor songs were presented to the females and songs
from a different area were presented to the females when they reached maturity:
References:
Baker, M. C. 1981. Early experience determines song dialect responsiveness
of female sparrows. Science 21: 819-821.
Index
Founder Effect Speciation
Founder effect speciation refers to a mechanism very similar to Allopatric
models of speciation. The primary difference relates to the size of the
founding population which colonizes some region on the very edge of a species
range. If this founding colony remains isolated for a long period of time
then speciation might occur. A key component of founder effect speciation
relates to the genetic material that is brought in by the initial colonists.
Because the number of colonists is very low, the sample of genetic material
is much reduced compared to the source population. In this case, the founding
population is expected to diverge from the source population from the outset.
Genetic drift facilitates divergence over the long term.
Index
Runaway Process, Sexual Selection, and Speciation
Recall the elements of Fisherian runaway process
and sexual selection. The essence of the theory is that a mutation for
female choosiness arises in a population and the females opt for males with
some kind of elaborate trait. If Fisherian runaway process is to play a
role in speciation mechanisms, then it must be related to the process by
which reproductive isolation occurs. How do the female descrimination traits
such as that seen in Gulls evolve? What about plumage variation seen in
other bird species? How is it that lizards have evolved such markedly divergent
head bobs.
The theory for runaway process as independently solved by Mark Kirkpatrick
and Russel Lande has a very interesting second phase.
In the first phase we saw that:
Fisherian runaway will lead to ever increasing escalation in the male
trait up to the point that natural selection begins to strongly act against
the handicap that the male trait imposes on the male. Once this point is
reached, the second phase of Fisherian runaway sexual selection takes hold.
The populations will change in frequency by the process
of what is refered to as genetic drift. Genetic drift is simply the
random processes that cause small fluctuations in the frequency of an allele
simply because of limited population size. By chance alone, the frequency
of alleles (for choosiness and the male trait) will fluctuate up and down
in frequency. In small populations the effect of genetic is expected to
be quite great compared to large populations. How small is small? Well generally
populations that are less than about 100 individuals. What might happen
is that certain alleles for choosiness might go to fixation in a small population,
or perhaps genes for the male trait.
Up to this point we have considered runaway selection and its operation
on a single male trait and a single mutation for choosiness. However, as
discussed above sexual selection operates on the whole
organism and females might choose on the basis of a number of male traits.
For example, females might choose on the basis of both tail length as well
as plumage coloration.
Now imagine a number of distinct sub-populations which are experiencing
genetic drift and going to fixation. Because genetic drift is an entirely
random process we would expect that each sub-population would fix for different
choosiness or male traits. When this occurs, we have evolved strong assortative
mating mechanisms that would lead to isolation.
Another scenario for such random effects is that the mutations
that lead to choosiness or the male trait might differ in each sub-population.
Fisherian runaway selection might lead to rapid evolution in each population
according to such initial differences. And the populations would be expected
to diverge rapidly in their mating preferences. The end result of runaway
process is reproductive isolation by mating preferences -- the key ingredient
for speciation.
Index
Back to the Syllabus