Back to the Syllabus
10. Dispersal, Navigation, and Migration
Barry Sinervo©1997
Index
Dispersal
Inbreeding avoidance
The Fitness Consequences of Inbreeding
Sex-biased Dispersal, Kin Avoidance, and Kin
Selection
Outbreeding Depression and Female Choice for
Natal Song
Extinction, Colonization Success and Dispersal
Navigation
Celestial Compasses
Bees and Polarized Light
Migration and Navigation in Birds
Zugenruhe
The Sun Compass of Birds
Stellar Navigation: "I got the sun in the
morning and the stars at night"
Planetarium Star Shifts
Learning or Instinct of the Stars?
Dispersal
Dispersal results in a tendency for animals that were once aggregated,
to become more widely distributed through movement away from aggregations.
The most common aggregrations found in nature are associated with the nest
site. Hatching juveniles or fledglings leave from a concentrated place,
and attempt to find a site that they might live in for most of the rest
of their life. Such long distance movement can be extreme in the case of
birds. Rare dispersal records based on banding at the nest and recovery
by other ornithologists have registered dispersal distances measured in
1000's of kilometers. Now this is not surprising given the flight capabilities
of birds. Dispersal of mammals is measured on the scale of tens of meters
and kilometers. Records for lizards place the upper limit on dispersal at
1.5 km. Given the length of a hatchling (2.5 cm), and the distance traveled,
1.5 km * 1000 m/km * 100 cm/m this would amount to 60,000 body lengths or
put into human body lengths (2m tall), the human equivalent of 120 km traveled
in the first week or two of life.
What could be worth travelling such inordinate distances?
There must at the very least be incredible energetic costs to such long
distance movement. In addition, there might be a disadvantage measured in
terms of survival. A moving animal may be at great risk of predation. We
will explore two theories that might explain why juveniles of most species
tend to disperse:
- Inbreeding avoidance
- Colonization of new habitat.
Index
Inbreeding Avoidence
The Fitness Consequences of Inbreeding
Inbreeding can be defined in many ways, but we generally think in terms
of the simplest definition which has to do with matings among related individuals.
However, as all humans ultimately share some related individuals (e.g.,
the proverbial adam and eve) at some point during their remote past, this
definition is not all that precise. A more precise definition that also
has some practical merit is to express inbreeding as the probability
of the two alleles on complementary chromosomes being identical by descent.
If these two copies of the allele (one from mother and one from father)
are identical by descent, then they must have arisen in the not-so-remote
past from a single strand of DNA. Calculations concerning the probability
of identity by descent has some practical merit because it can be used to
calculate the risk that a a deleterious recessive mutation might show up
in an inbred individual. If an individual has two identical copies of a
deleterious rececessive mutation, that individual will express a
trait that in some way lowers fitness, or in the worst case of a deleterious
recessive lethal, will cause death.
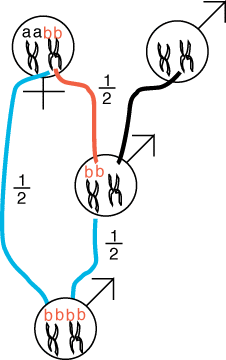 The easiest way to get two
genes together that are identical by descent is from a consanguinous
mating, or a mating among close relatives. Consider progeny fertilized from
a mating between a mother and son and try to track the alleles coming from
the mother. We will track one allele, bb, from
mother to son and from the mating of mother and son which we will call an
oedipal mating. (Recall that each pair of chromosomes posseses two sister
chromatids). The allele b is located on one
of the mother's two chromosome pairs.
The easiest way to get two
genes together that are identical by descent is from a consanguinous
mating, or a mating among close relatives. Consider progeny fertilized from
a mating between a mother and son and try to track the alleles coming from
the mother. We will track one allele, bb, from
mother to son and from the mating of mother and son which we will call an
oedipal mating. (Recall that each pair of chromosomes posseses two sister
chromatids). The allele b is located on one
of the mother's two chromosome pairs.
The son may receive one copy of allele b
with probability of 1/2 and this transmission is denoted by the red
line.
The blue lines denote the 1/2 probability
that the mother will contribute the b to a
progeny sired by her son, who also contributes allele b
with probability 1/2 via the second blue line.
Because all events are mutually independent, we multiply all of the probabilities
to get the net probability that a child produced by an oedipal mating possesses
two bb alleles:
1/2 * 1/2 * 1/2 = 1/8 (eqn 1).
Note that, the "a" copy could just as easily been transmitted
to the son in the first place so the total probability that the alleles
in the oedipal child are identical by descent can be achieved two ways,
either with allele "a" or with allele b
and the probability of inbreeding from an oedipal mating is:
2 * 1/2 * 1/2 * 1/2 = 1/4 (eqn 2).
Of course, there is a 3/4 probability that the oedipal child is not identical
by descent. Let us consider a few special cases to assess the fitness consequences
of such inbreeding. If mom was a carrier for a single deleterious recessive
allele, b, then the probability that a son
produced from an oedipal mating would die is given by the probability that
a single allele is identical by descent = 1/8. (Note that mom has one good copy in the form of allele
"a".
If the mom happened to carry an allele that was deleterious on each of
two different genetic loci we would multiply this value by 2 to get the
probability of a progeny from an oedopil mating getting at least one such
double whammy = 2 * 1/8 = 1/4. Because each an individual may possess several deleterious
recessive alleles, matings among related individuals typically produces
inbreeding depression or a reduction in fitness that arises from
identity by descent. Such alleles do not have to be lethal, rather
the combination of several mildly deleterious alleles can have a
dramatic impact on fitness.
Index
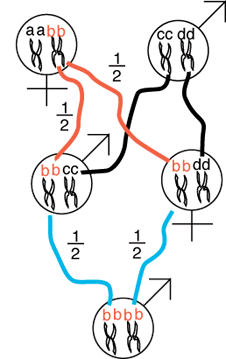 As a second example, let
us consider the possibility of a sib-sib mating. We now have to keep track
of the fathers alleles because a grandchild can become inbred from one of
the mom's alleles or one of the the father's alleles. Again, we will track
one allele, bb, from mother to son and from
the mother to daughter. At the end we will then multiply the probability
of a single allele being identical by descent by four (all possible alleles
that might be identical by descent) to determine the overall probability
that any allele is identical by descent.
As a second example, let
us consider the possibility of a sib-sib mating. We now have to keep track
of the fathers alleles because a grandchild can become inbred from one of
the mom's alleles or one of the the father's alleles. Again, we will track
one allele, bb, from mother to son and from
the mother to daughter. At the end we will then multiply the probability
of a single allele being identical by descent by four (all possible alleles
that might be identical by descent) to determine the overall probability
that any allele is identical by descent.
The mother gives the b allele to her son
with probability of 1/2 and likewise the daughter receives the b
allele with probability of 1/2 (both denoted by the red
lines). Given that the daughter received the b
allele, the daughter passes the allele on to her own progeny with propbability
1/2 (blue line). Likewise, given that the son
has the b allele he passes it on to his progeny
with probability of 1/2.
The, probability that the grandchild received two copies of the b allele
is given by:
1/2 * 1/2 * 1/2 * 1/2 = 1/16.
Finally, we realize that the grandchild could just as well have received
the a, b, c, or d alleles by the same routes with the same probabilities
so the probability of any two alleles at a single locus in the same individual
being identical by descent is given by:
4 * 1/2 * 1/2 * 1/2 * 1/2 = 1/4.
The same logic can be used to compute the probability of two cousins
yielding a child that has genes identical by descent, though the length
of the paths are a little bit longer. Indeed, the logic used above can be
used to compute the probability of inbreeding or identity by descent
for any consanguinous mating. The inbreeding in any set of pedigrees
can be computed from similar path diagrams that chart the genealogical
relationships among individuals.
Index
Sex-biased Dispersal, Kin Avoidance and Kin
Selection
The fitness costs arising from such inbreeding mechanisms have lead various
authors to propose that patterns of sex-biased dispersal are a kin avoidance
mechanism. The tendency for offspring to breed in their natal home range
is referred to as philopatry. Because dispersal per se entails costs,
both sexes of progeny need not disperse to dramatically lower the probability
of inbreeding. If either the female progeny disperses or the male progeny
disperses, the risk of inbreeding is lowered dramatically because one of
the most pernicious sources of high inbreeding (sib-sib is eliminated, still
have to worry about father-daughter). Most mammals have male-biased offspring
dispersal, and the female progeny tend to be philopatric. For example, Kay
Holekamp has found that female ground squirrels tend to remain near their
natal nest and males disperse several hundred meters away.
The tendency for dispersal in one sex cannot be explained in terms of
kin avoidence, but ideas of kin selection must be used to explain
why female progeny remain. If female's get assistance from their mother
or from sisters, then they would be likely to be philopatric.
Kin selection appears to be related to levels of philopatry in other
mammals and the dispersing sex is the sex least likely to benefit from kin
selection. For example, in cheetahs, if a female cheetah produces two or
more sons, those sons are likely to form a kin group and defend a territory
cooperatively. The brothers will even participate in cooperative breeding.
It takes a few males to adequately defend a territory against other males.
Conversely a solitary male lion usually heads a pride and there is no
advantage to being philopatric in lions. Male lions disperse, whereas females
remain with the pride. To further avoid inbreeding between a father and
his daughters, many male lions disperse and attempt to take over another
pride of lions when his daughters begin maturing. This is fairly common
in other groups were the possibility of father-daughter matings arises.
What evidence is there that inbreeding can be costly in nature?
Very little evidence is available that directly links inbreeding depression
to lack of dispersal in wild populations, but Packer (1979) reported anectodal
information on a male baboon that failed to disperse at sexual maturity.
He produced low surviving progeny compared to outbred males in the baboon
troop.
Information on inbreeding depression per se is available for cases in
which researchers have drawn up pedigrees of the sort shown above. An elegant study of such pedigrees for sparrows
on Mandarte Island (XXX and Smith, 1987) illustrates real costs to inbreeding.
However, in the case of the song sparrows, the effects of reduced survival
in inbred individuals were most strongly manifest during periods of environmental
stress. Those individuals with a higher inbreeding coefficient (e.g., the
product of one or more consanguinous matings based on the pedigree computations)
were more likely to die during a famine than individuals with a low inbreeding
coefficient.
Index
Outbreeding Depression
While consanguinous matings generally result in inbreeding depression,
it is does not follow that an animal sould find a mate that has a completely
different genetic background. Outbreeding depression is the tendency
to show reduced fitness in progeny that are the product of two very different
genetic backgrounds. There may be an optimum level of outbreeding. Indeed
the optimal dispersal distance may reflect such outbreeding depression.
Go too far and you end up in the company of very different genetic mates.
Outbreeding depression is thought to arise from epistatic
interactions among loci. A minor amount of incompatibility among a suite
of genes involved in some sort of enzyme cascade may lower fitness.
Female Choice for Natal Song
Outbreeding depression may give rise to a behavioral block to breeding
that is similar to species recognition mechanisms. For example, many birds
are only sexually responsive to mates from their own natal areas.
The following experiment illustrates that female birds learn songs of appropriate
mates in their youth and then choose rather srupulously regarding males
that sing the right song.
Baker first used tudor males to teach young birds a song
dialect:

Songs similar to the tudor songs were presented to the
females and songs from a different area were presented to the females when
they reached maturity:
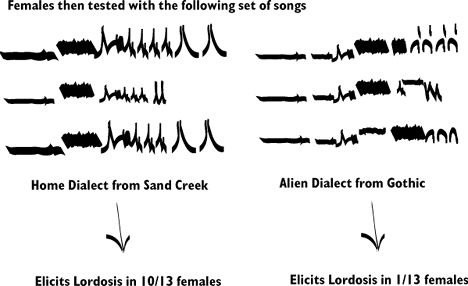
Baker, found that female white-crowned sparrows responded
positively to male courtship songs at a higher rate if the song was derived
from their home dialect compared to an alien dialect.
The key feature of this study demonstartes that birds learned
the song during the nestling phase. In addition, it is possible to fool
females into preferring a truly "non-natal" song, or one from
a region other than their true source population. By playing them a song
that is from a different source population during the critical song learning
period. For example, if tudor songs had been from Gothic the females should
have preferred Gothic males at maturity and rejected Sand Creek Songs.
References:
Baker, M. C. 1981. Early experience determines song dialect responsiveness
of female sparrows. Science 21: 819-821.
Index
Extinction, Colonization Success and Dispersal
 The advantages and disadvantages of dispersal strategies
are not limited to genetic costs of inbreeding. Consider, a meta-population
in which many sub-populations are found loosely connected and each sub-population
is relatively small. Moreover, each sub-population has a modest probability
of local extinction where all members of the population might be
eliminated by some random environmental disaster.
The advantages and disadvantages of dispersal strategies
are not limited to genetic costs of inbreeding. Consider, a meta-population
in which many sub-populations are found loosely connected and each sub-population
is relatively small. Moreover, each sub-population has a modest probability
of local extinction where all members of the population might be
eliminated by some random environmental disaster.
In such a case, selection will favor individuals that disperse from occupied
habitats to those parts of the habitat where a local extinction has occurred.
Such disperser genotypes do not have any inside information about
the location of "open" habitat, but if a lucky disperser happens
upon such unoccupied habitat their reproductive rate will be quite high
compared to those individuals found in the occupied and crowded habitat.
A disperser landing in an occupied area has low success. Theory predicts
that the dispersal tendency will be quite common even with a low frequency
of local extinctions of a sub-populations.
The Eastern newt, Notophthamalus viridescens appears to be a candidate
for an extreme version of this model. The extinction of newts in a pond
is a very likely event if the adults are not successful in producing metamorphlings
across a string of bad years (Gill 1980). Adults breed in temporary ponds,
and the adults return to the same pond year after year. The reproductive
success of newts in any one pond is extremely low, and the newts go extinct
in ponds quite frequently. This species of newt has evolved a dispersing
phase to its life cycle called the "red eft" that wanders
around the woods moving great distances over the course of several years.
The eft eventually settles in a pond, and if it happens to settle in a "near
empty" pond, with at least one mate, the two metamorphosed newts will
likely colonize the pond and have high success if they get lucky. The newts
appear to be a case in which an obligate dispersal phase has
been favored. All offspring turn into an efts and disperse.
Other species of amphibians illustrate that two or more dispersal types
may coexist in the same population. In this case, the dispersal tendency
is facultative and triggered by density of conspecifics. Semlitch
has identified genetic predispositions for two kinds of salamanders:
- metamorphosing from a larva to terrestrial adult, which may or may
not come back to breed in the same pond,
- or not metamorphosing and remaining in the pond and breeding in the
larval body type.
The type of salamander that does not metamorphose, also does not disperse
and is known as a paedomorphic salamander. Because paedomorphic salamanders
remain in the water their entire life, they are susceptible to a large-scale
drying event, because they have lost the capacity to metamorphose. However,
because paedomorphic avoid the costs of dispersal, they grow to a larger
size at maturity, and have a competitive advantage over the form that has
the typical amphibian metamorphosis and dispersal phase. Even if the paedomorphic
form goes extinct and the pond is recolonized by the typical form, new paedomorphic
mutants are probably arising all the time. The evolutionary switch or mutation
that gives rise to new paedomorphic forms from the typical forms is very
simple. All that is entailed is a downregulation in the genes that cause
metamorphosis from aquatic larvae to terrestrial adult (Thyroxine production
and regulation). The other more typical form possesses fully functional
genes and metamorphoses under most conditions. This form is much more likely
to colonize the newly extinct ponds. In the long term, it has an advantage
over the paedomorphic form because all ponds will eventually dry up and
the population goes extinct. As new mutations can always arise and invade
it is an excellent example of a system that has no Evolutionary
Stable Strategy. Both forms coexist with one periodically outcompeting
the other, yet neither is a longterm winner.
Index
Migration, Orientation, and Navigation
Dispersal is typically associated with a once in a lifetime event
-- movement to a new habitat. During the dispersal event, it is thought
that animals might make a random movement, and then settle once suitable
habitat is found. Once the animal has settled in a suitable patch, it must
move around its local universe and not get lost.
We will explore how bees and birds use visual cues such as the sun or
the starts to orient. We will consider magnetic navigation in birds. Orientation
is the use of external cues to move about the environment. Orientation can
come about by fixing a position and then using cues to determine the appropriate
direction to move in. Navigation involves a little more sophistication
in that a map sense (a sense of where you are) in addition to a compass
sense (a sense of the appropriate direction in which to move) are a requirement
for a complete navigational system. Many studies have demonstrated orientation,
but few have demonstrated that animals have a map sense or a sense of where
they are.
Migration is distinct from dispersal in that migratory animals
typically move from one geographic region to another,without using the intervening
habitat. The movements are associated with a particular season, and invariably
tied into the reproductive cycle. A key issue underlying migration is how
animals navigate the huge distances or orient to natal areas. Underlying
most migrations is a sense of time so that the appropriate season
for migration is used. Migration is classically associated with birds. However,
many other groups undertake long migratory movements:
- insects (e.g., Monarch butterflies),
- mammals (caribou, wildebeests, whales),
amphibians (e.g., newts to their natal pond),
- reptiles (e.g., sea turtles),
- fish (salmon, eels).
Celestial Compasses
A pervasive feature in the environment that animals could use for orientation
is the sky itself. The sun rises at specific times during the day, and provided
that an animal has a sense of time, the relative position of the sun could
be used to determine direction. However, cloudy skies limit the use of the
sun as a source of orientation information. What remains intact on even
the cloudiest of days is polarized light. Light rays entering the atmostphere
become scattered. Owing to some fancy physics, this creates polarized light.
The sky in the direction of the sun is less strongly polarized than the
sun in the other half of the sky which is maximally poloarized -- even on
a partly cloudy day. Of course a cloudy day does obscure the polarization
quite a bit, but it remains a steady albeit attenuated environmental cue
that could be used if organisms could detect polarized light.
What is polarized light?
 Tough to visualize. Well imagine you had eyeballs sensitive
to polarized light and you looked at the sun. The light from the sun is
coming from all directions and is not all that polarized, so the sun would
look dark (solar sky). As you turn your head away from the sun and look
90 degrees away, the polarized light gets brighter and brighter. Exactly
90 degrees away is the brightest for polarized light because a lot of light
rays are heading perpendicular to your field of view. As you continue turning
your head, further away from the sun, the polarized light gets dimmer and
dimmer, until it is dark again when you are looking away from the sun (anti-solar
sky). This is because light is again heading in away in all directions compared
to when you are looking 90 degrees from the sun when it is coming from the
sun perpendicular to your field of view.
Tough to visualize. Well imagine you had eyeballs sensitive
to polarized light and you looked at the sun. The light from the sun is
coming from all directions and is not all that polarized, so the sun would
look dark (solar sky). As you turn your head away from the sun and look
90 degrees away, the polarized light gets brighter and brighter. Exactly
90 degrees away is the brightest for polarized light because a lot of light
rays are heading perpendicular to your field of view. As you continue turning
your head, further away from the sun, the polarized light gets dimmer and
dimmer, until it is dark again when you are looking away from the sun (anti-solar
sky). This is because light is again heading in away in all directions compared
to when you are looking 90 degrees from the sun when it is coming from the
sun perpendicular to your field of view.
Index
Bees and Polarized Light
 The celestial compass of bees was discovered
by somewhat indirect means. Bees go out on foraging flights, find food,
then fly back to the hive to communicate to the other workers the source
of the food.
The celestial compass of bees was discovered
by somewhat indirect means. Bees go out on foraging flights, find food,
then fly back to the hive to communicate to the other workers the source
of the food.
The bees use a waggle dance to indicate the direction of the nectar source.
The sun provides a very constant reference in the sky for the bees to
direct other workers. The bee ends its circuit around the loop with a waggle,
and the distinctive waggle part of the dance lines up with a vector that
leads directly to the nectar source. The distance from the hive is proportional
to the number of waggles in the dance.
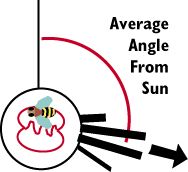 Karl von Frisch decoded the language of bees
using some clever manipulations. The tendency for bees to point in the correct
direction of the nectar supply can be plotted on a circle. The height of
the histogram is proportional to the number of worker bees thatchoose that
vector (i.e., typically close to the actual angle of the plant from the
sun if they are correct). Few workers are far off the actual angle.
Karl von Frisch decoded the language of bees
using some clever manipulations. The tendency for bees to point in the correct
direction of the nectar supply can be plotted on a circle. The height of
the histogram is proportional to the number of worker bees thatchoose that
vector (i.e., typically close to the actual angle of the plant from the
sun if they are correct). Few workers are far off the actual angle.
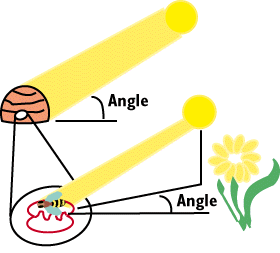
In the diagram to the right, the Average Angle of the Worker Bees dance
indicates that food is located approximately 100 degrees to the right of
the sun. A worker would fly at a bearing of 100 degrees to the sun and come
upon the source of food that the scout bee had found. It appears that the
worker is using the sun a reference. In actuality the bee is using polarized
light as a reference. The omatidia of the bee contain special photoreceptors
that are sensitive to polarized light in ultraviolet wavelengths.
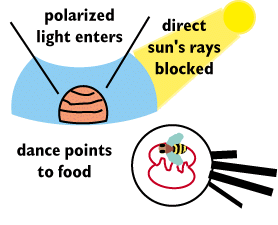 Wehner and Rossel discovered the workings of the bees
celestial compass by manipulate the light coming into the hive while the
bee did its waggle dance.
Wehner and Rossel discovered the workings of the bees
celestial compass by manipulate the light coming into the hive while the
bee did its waggle dance.
They showed that the sun's rays were not necessary by putting up a screen
that eliminated the sun's direct rays from entering the hive, while permitting
the skys polarized light to reach the hive through an opening in the top
of screen.
Worker bees will display the correct direction to nectar even though
the sun is not visible.
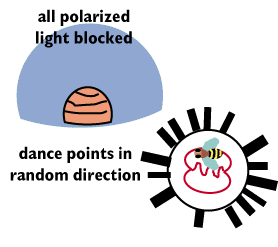 If you block out all polarized light using a plexiglass
bubble, you eliminate the cue that the bees use too orient themselves. The
plexiglass absorbs polarized light in the ultraviolet wavelengths.
If you block out all polarized light using a plexiglass
bubble, you eliminate the cue that the bees use too orient themselves. The
plexiglass absorbs polarized light in the ultraviolet wavelengths.
A bee that has come back from its scouting knows the correct angle, but
without the sun to guide it, the worker cannot tell which direction it should
end its waggle so as to direct the workers. The data for a number of workers
shows a very random pattern heading off in every direction compared to the
consistent directions shown above.
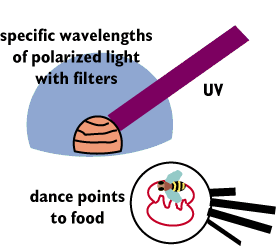 Finally, one can let in just certain wavelenghts of
polarized light using filters. In such experiments, the researches keep
altering the wavelengths until the hit upon the exact wavelengths that give
the bees the external information it needs to orient their dances correctly.
Many wavelengths of light give a random orientation to the dance pattern.
However, light in very specific uv wavelengths appears to trigger the polarized
light sensitive regions of the omatidia and allows for correct orientation.
The bee is "blind" to such directional information even if the
light is correctly polarized, but in the wrong wavelengths.
Finally, one can let in just certain wavelenghts of
polarized light using filters. In such experiments, the researches keep
altering the wavelengths until the hit upon the exact wavelengths that give
the bees the external information it needs to orient their dances correctly.
Many wavelengths of light give a random orientation to the dance pattern.
However, light in very specific uv wavelengths appears to trigger the polarized
light sensitive regions of the omatidia and allows for correct orientation.
The bee is "blind" to such directional information even if the
light is correctly polarized, but in the wrong wavelengths.
Index
Migration and Navigation
Zugenruhe
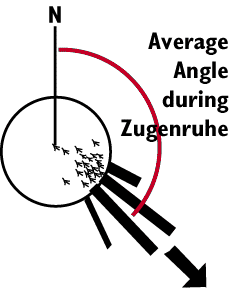 Birds use the sun as a simple compass. How do we know
that that they use it for orientation? Like the bees, the birds indicate
the direction that they wish to fly by a well known phenomenon called the
zugenruhe or nighttime restlessness. Zugenruhe refers
to the sharp jump in nighttime activity that occurs at the onset of the
migratory response. Because birds migrate at night, but are not normally
active at night during other times of the year, students of navigation have
used birds as a model system for understanding directional orientation.
Birds use the sun as a simple compass. How do we know
that that they use it for orientation? Like the bees, the birds indicate
the direction that they wish to fly by a well known phenomenon called the
zugenruhe or nighttime restlessness. Zugenruhe refers
to the sharp jump in nighttime activity that occurs at the onset of the
migratory response. Because birds migrate at night, but are not normally
active at night during other times of the year, students of navigation have
used birds as a model system for understanding directional orientation.
Another aspect of zugenruhe is that birds orient strongly in the
direction in which they are to migrate. A clever apparatus consisting of
a cone shaped cage with a mesh top (Emlen 1970), and an inkwell at the bottom
has been developed to record the directionality of the zugenruhe.
The birds hop up and down trying to take flight, and end up leaving a complete
record of each hop, as well as the direction of the hopping relative to
magnetic north.
Index
The Sun Compass of Birds
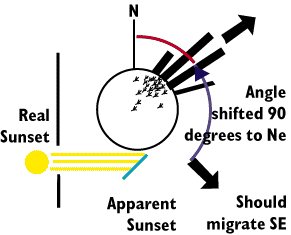 Let's consider Starlings that have a normal migration
route that is in a SE direction. It is easy to shift the direction of zugenruhe
by using mirrors to alter the true direction of the sun at sunset by 90
degrees clockwise or counterclockwise. Starlings in aviaries that see such
a frameshifted sun end up leaving tracks that are 90 degree frame shifted
in the appropriate direction.
Let's consider Starlings that have a normal migration
route that is in a SE direction. It is easy to shift the direction of zugenruhe
by using mirrors to alter the true direction of the sun at sunset by 90
degrees clockwise or counterclockwise. Starlings in aviaries that see such
a frameshifted sun end up leaving tracks that are 90 degree frame shifted
in the appropriate direction.
Researchers hypothesize that the birds take a bearing on the sun at sunset
and then use dead reckoning to fix on a correct direction until sunrise
at which point a new bearing can be used. Regardless of the mechanism, it
is clear that the birds use the sun as a Celestial compass much like bees
used polarized light as a Celestial Compass.
Overcast Skies
 If the skies are overcast, then the birds have a random
orientation to their zugenruhe movements. This suggest that the sun is necessary
to at least get a quick fix on or the birds do not have a directional cue
to initiate their dead reckoning during the night.
If the skies are overcast, then the birds have a random
orientation to their zugenruhe movements. This suggest that the sun is necessary
to at least get a quick fix on or the birds do not have a directional cue
to initiate their dead reckoning during the night.
If birds do not have the sun, what other cues might they use? Some have
suggested the weather. Winds are very predicable at certain altitudes and
a stiff cross wind might be used to aid the bird during periods of no sun
or to further refine orientation at night. During the day, birds undoubtedly
use large landmarks like the Mississippi River, and such bodies of water
would also be available at night via the reflection of the moon.
What other cues might birds use at night?
Index
Stellar Navigation: "I got the sun in
the morning and the moon and stars at night"
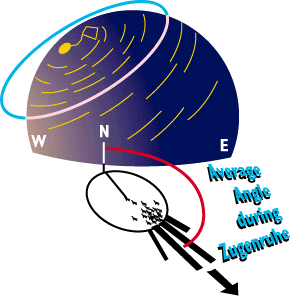 A little primer on stellar navigation.
A little primer on stellar navigation.
Some birds are real stargazers, and can use the position of the stars
to orient.
Most of us have looked up in the night sky and you may have noticed that
the constellations move during the course of the night. This apparent motion
is caused by our own earth's rotation, and polaris happens to be located
directly along the earth's north-south axis. As the earth rotates, polaris
does not move, but the other starts appear to rotate around polaris which
is the handle-tip to the little dipper.
Planetarium Star Shifts
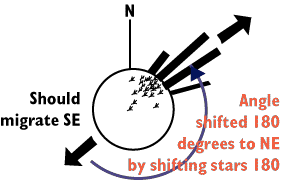 If birds are set up in a planetarium all of the stars
can be projected on the dome like ceiling, and the rotational position of
the stars can be easily manipulated by the machine that projects the stars
on the ceiling. The orientation angle during zugenruhe can be used
during such manipulations of stellar cues to determine what information
birds might be using at night.
If birds are set up in a planetarium all of the stars
can be projected on the dome like ceiling, and the rotational position of
the stars can be easily manipulated by the machine that projects the stars
on the ceiling. The orientation angle during zugenruhe can be used
during such manipulations of stellar cues to determine what information
birds might be using at night.
First, Sauer simply manipulated the stars images projected on the dome
of the planetarium. He rotated the stars by 180 degrees. The star shift
is not just a rotation of the stars (which birds could use as a clock, much
like we use the sun during the day), but rather, the orientation of polaris
and all stars around polaris are flipped into the souther sky. As predicted,
he rotated the direction of the Zugenruhe by exactly 180 degrees.
Birds use some kind of view of the stars to orient their zugenruhe.
Are birds born with a sense of the stars or do they learn it?
Index
Learning or Instinct of the Stars?
Emlen conducted an experiment on the early experiences of Indigo buntings
with a star map. He set up two groups of chicks that were reared under the
following conditions:
- chicks reared in a room with a diffuse light at night
- chicks reared with a normal star map that rotates around
the north star polaris
- chicks reared with a abnormal star map that rotates around
Betelguese which is a star on the southern constellation of Orion.
All the birds were then tested with a normal star pattern (even those
reared under the baleful eye of Betelgeuse).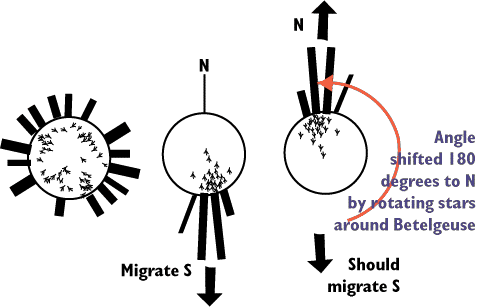
As predicted, birds without a star chart did not learn
the appropriate cues and showed no orientation.
Those birds with a reversed experience of the stars also
showed a reversed Zugenruhe relative to the control birds.
Birds learn the star map in their youth and use this information to orient
their Zugenruhe, which is a good thing, as the earth wobbles and thus the
star map is not constant, but it changes every 13,000 years or so.
Index
Back to the Syllabus