


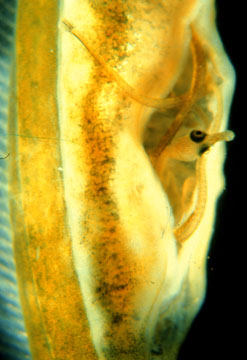
male brood pouch
copulation
full brood pouch
birth
fry and father
The Ecology and Evolution of Mating Systems
Sticklebacks, Pipefish, and Seahorses: Polygyny, Polyandry, and Monogamy
Polygyny, Monogamy and Sperm Competition in the Rock-paper-scissors Game
Multiple Paternity and "Monogamy" in Birds
The Evolution of Parental Investment
Monogamy, Polygyny and Offspring Quality in Pied Flycatchers
Sexual selection is the driving force that underlies the evolution of male competition and female choice, but what ecological factors contribute to one species having highly ornamented males and another having no differentiation between the sexes. Not all species show strong sexual dimorphisms. In fact many species of monogamous birds have males and females are very similar in outward appearance and it is impossible to sex them by external features alone. Clearly sexual selection does not take hold and lead to a runaway or good genes process in all species on the planet.
We will explore a diversity of mating systems in an attempt to understand why some species are monogamous and others are polygamous.
Monogamous species of males and females pair for the breeding season. In some cases, the individuals may pair for life.
In polygynous species, males have multiple female partners in a single breeding season. In some cases a male may control a large group of females in other cases the polygyny may paractive sequential polygyny, in which a male visits with successive females.
The flip side of polygynous is a polyandrous mating system in which females have multiple male partners.
There are also subcategories of polygynous and polyandrous mating systems that are defined in terms of whether the mating systems are resource based or non-resource based. In a resource based polygynous mating system a male defends some resource that might sustain several females. In a non-resource based mating system like a lek, females aggregate in regions with sole purpose of choosing a mate. Males aggregate in these locations and display in order to gain access to females.
Sexual selection theory and the relatively low parental investment that males make in offspring would suggest that polygynous mating systems should predominate in the animal kingdom. The real puzzle is posed by the following question:
In a classic paper, Emlen and Oring (1977), suggested that the ecology of the organism may not permit males to have more than one partner. Females may be so widely distributed that males cannot monopolize more than one female.
The male is not the only individual making such choices, for the female plays an active role in the decision making regarding settling with a male that already has a female or searching for a male with an unoccupied territory. Gordon Orians (1969) advanced the Polygyny threshold model as the basis by which females decide to enter into a monogamous versus polygynous mating.
Alatalo and Lundberg have shown tha female pied flycatchers that are mongamous have higher reproductive success compared to females that settle with a polygynous male. In addition, the primary female of a polygynous pair (first arrival), has higher reproductive success compared to the secondary mate (second arrival).
Emlen, S. and L. Oring. 1977. Ecology, sexual selection, and the evolution of mating systems. Science 197: 215-223.
Orians, G. 1969. On the evolution of mating systems in birds and mammals. American Naturalist 103: 589-603.
In the evolution of a resource based mating system, females end up settling on a male's territory or depositing eggs on a male's territory because of the quality of his territory. It is thought that the males with the best condition, bright badges of status, the sexual-selected Vunderkind of the animal kingdom end up controlling what are called hot spots where females aggregate and spend most of the breeding season.
If resources are more diffuse, then males cannot practice a resource-based polygyny. It is thought that another system might evolve from a resource-based system:
When this state is reached, it is termed a Lek, a place where males display in groups and females choose.
Females Come around and pick from the available males. The paradox of the lek is that they usually mate with one male. Yes all females line up and mate with one male.
The real paradox, is why the other males bother to come to the lek at all if only a single male is going to get copulated. These males would do far better to go in search of females becaue they would at least get one or two copulations and not up end as big losers.
Many species form leks: Grouse, manikins, fish (pipefish females), insects, marine iguanas, etc. It is nearly a winner take all form of mating. There are big winners and big losers.
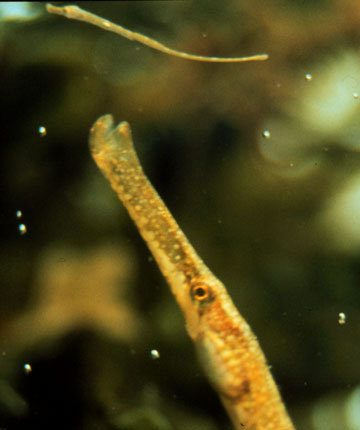
Sticklebacks, pipefish, and seahorses belong to the same family of fish. This group is noteworthy in the animal kingdom for the evolution of highly-advanced male care. This system is also illuminating with regards to the role of ecology in governing the evolution of mating systems.
The evolutionary relationships among these three groups is roughy as follows:
Sticklebacks -> Pipefish -> Seahorses
All groups have male care, but the fish are found in distinctly different habitats have a profound effect on the form of mating system that runs from:
Polygyny -> Polyandry -> Monogamy
These fish are found in the following habitats:
Estuaries -> Eel grass -> Diffuse on Reefs
While most species of sticklebacks are found in very dense aggregations, females are still limiting because a male can mate with more than one female and care for her eggs in his nest. Males are polygynous and strongly selected for the red ornament.
In the case of pipefish, males are now limited by the size of the brood
pouch, and females are found in excess relative to the number of non-gravid
males. Females and males are found in dense aggregations in eel grass beds,
and this leads to strong local mate competition among the females. The females
that are the largest and most ornamented are victors in this competition.
The pipefish are also a largely temperate species, and their breeding takes
place during a limited time of the year. Thus there are many males available,
but only for a short time, as females scramble to visit as many males as
they can.
| |
|
|
|
|
male brood pouch |
copulation |
full brood pouch |
birth |
fry and father |
Males are now the choosy sex owing to an over abundance of females. Rosenqvuist assessed the importance of the female ornament in male choice by presenting females of different size and who varied in the degree of ornament development to males . Overwhelmingly, males preferred females that were larger and more ornamented, the key conditions necessary for the evolution of even more elaborate size and ornamentation in female pipefish. Below you will find additional examples of sexually selected pipefish and the elaboration of female ornaments.
 |
 |
Nerophis Ophidon male |
Nerophis Ophidon female |
|
Syngnathus nigra |
The situation for seahorses is similar in terms of the elaborate male care, but seahorses differ from pipefish in one important ecological factor. Whereas pipefish are found in dense aggregations in eel grass habitats, nearly all species of seahorses are very diffusely distributed around reefs or in sargassum. The density of males is in fact so low that once a female has found a male, she would probably have higher fitness defending the male against other females and entering into a monogamous relationship, than if she were to go in search of another male. The likelihood of finding an unoccupied male is rather remote. The probelm is compounded, in that unlike pipefish, which breed in a very short time window, seahorses have a long breeding season. Females remain with the male during the entire breeding season, ovipositing her eggs in his brood pouch with each successive birth. With the evolution of monogamous mating system, one finds far less sexual selection and seahorse males and females are very similar in morphology. Many seahorses are selected for crypsis, as ornaments would be debilitating to survival during a long breeding season.
|
|
Solenostomus paradoxus |
H. breviceps |
I will go over the web page describing the mating system of side-blotched lizards and place special emphasis on the role of territoriality in defining the three male strategies.
 the largely
monogamous mate-guarding Blues
the largely
monogamous mate-guarding BluesIn our visit back to this unusual mating system, I will describe some new DNA paternity results from our lab that document some very unusual aspects relating to sperm competition.
Many birds are essentially monogamous and mate for life. Before the advent of DNA paternity, most scientists truly believed that monogamous animals were monogamous. After DNA paternity it was quickly realized that many monogamous species participate in what are referred to as extra pair copulations or EPCs. Nearly every supposed mongamous species participates in EPCs. It is not just the males that go and solicit copulations from neighboring females. Some females go in search of other males and solicit copulations.
For males the question is obvious, a male that solicits additional copulations gains a direct increase in fitness.
For female the question of EPCs is a little more difficult to answer.
A single male is capable of fertilizing all of a females clutch, so why should she go and solicit additional copulations. There might be costs to such search behavior. In addition, in a monogamous species in which the male and female provide care to the young, a male that detects such extra-pair activities, might be expected to abandon the female because the certainty of his paternity comes into question. Why should he rear offspring that are not related to him?
The most important reason cited for extra-pair or multiple partners is the possibility that a female is getting extra sperm to ensure that all her eggs are fertilized. If her choice of mate turned out to be infertile, then she would be wasting a lot of energy. Thus, fertility assurance would provide a direct benefit to the female.
There is evidence accumulating that many species (not just monogamous species) participate in multiple matings to ensure that all embryos are fertilized. The European Adder studied by Tom Maddsen and Matts Ollson is a case study. Females adders that participate in multiple matings have a lower risk of producing still born offspring.
A few researchers have even suggested that sperm competition per se could lead to the "fittest" sperm reaching the egg. You are used to thinking of the sperm of animals as an extension of the male, not really a separate organism. Well, haploid animal sperm could be thought of as a separate phase of an animal life cycle -- a self-contained organism unto itself. The sperm is free-swimming and must reach the egg before it dies. The sperm of highest quality might also be carrying high quality genes to the egg. Thus a female that participates in multiple matings might be setting up a sperm competition in which the sperm from many males compete and the best sperm wins the race.
The female not only gets offspring that might be of superior genetic quality, but she also gets offspring that are quite variable. Half-sibs have greater genetic diversity than full sibs. Such genetic diversity might be favored by selection. Imagine an organism that is found in a fairly variable environment in which not all the offspring from a female are going to survive because they do not have the necessary attributes that are dictated by the environment that they face. A female that produces highly variable offspring might be expected to have an advantage over a female that produces more uniform offspring. At least one of the offspring from the highly variable clutch will survive, whereas none from the less variable clutch will survive.
Why should animals produce males? Let us consider a concrete example -- the parthenogenetic whiptail lizards of the desert southwest. Scientists have found entire populations of these lizards that have evolved to produce fertile eggs without the benefit of sperm. Females reproduce in a way that literally leads to clone production. There is absolutely no genetic variability among her offspring and she only produces daughters. Now, consider the population growth rate of such a clone in competition with a normal sexual population. The clone grows twice as fast, because the normal sexual population in which females "waste" their energy producing sons that add nothing to the overall growth rate of the poulation. Only daughters produce offspring -- males fertilize. Such arguments have lead us to question the very existence of sex because the planet earth should be populated with only clonal species -- they reproduce with much greater efficiency. The cost of sex is the reflected in a decreased population growth rate (1/2) in a sexual population related to an asexual population.
Muller a nobel prize winning geneticist came up the following argument regarding clones.
All clones accumulate mutations.
The mutational load in an asexual lineage cannot decrease below that in the least-loaded clones, but the load can increase in all clones as they acquire new mutations. The least-loaded clone has the fewest deleterious mutations.
Muller's ratchet clicks with each new mutation, and with each click of the ratchet the overall fitness of the population declines ever so slightly. To be sure, this leads to an advantage in the least-loaded clone, but eventually that lineage will accumulate mutations or be wiped out by some unforeseen disaster.
Natural selection can purge the mutations from the population. Sex has evolved as a way to create diversity, recombination, that allows exsiting variation to be generated by scrambling the material between male and female parents. In addition, sex generates the raw material that natural selection can act upon, leading to steady "improvements" in the species.
Our own Bill Rice has argued that sex produces the rare progeny not at all like the parents and thus parasites can't infect their own offspring.
Asexuals produce identical progeny which are highly susceptible to infection.
Given the prevalence of parasites in most animals, sex would lead to a mechanism that would keep some offspring parasite free, at least until the parasite evolves resistance to whatever kept it from infecting the offspring in the first place. Sex is continually required to generate new variation because of this coevolutionary arms race between parasites and their hosts. As the host evolves new resistance, the parasites evolve mutations to crack the resistance.
Parental Care carries several distinct benefits, and it is curious why it is not pervasive in the animal kingdom. An organisms that gives its offspring a boost would be at a tremendous advantage over another that provided less parental care. Parental care can even include extra energy added to eggs when they are laid in organisms without parental care. This strictly material provisioning benefits the offspring tremendously. Bigger is usually better. Thus, the arguments described below are equally applicable to animals with and without parental care.
David Lack reasoned that parent care or provisioning is a relatively expensive proposition. There is a cost of reproduction that forms some sort of limit on how much an animal can do to provide care to its offspring. Lack also came up with an experimental paradigm for testing whether such costs of reproduction exist in birds.
Lack manipulated clutch size rand he reasoned that the parental bird would work harder to fledge the extra young and the extra cost of parental care would lead to increased mortality to the next breeding season. Since Lack's time, researchers have documented such costs of reproduction. They have also found that survival costs were not the only way energy limitation might manifest itself as a drop in fitness. In experiments with some species, the researchers found that they did not have a dramatic impact on the survival of the parents. Rather, they found that the size of fledglings or the number of fledglings that survived was reduced in nests that received extra eggs compared to control nests. Fledgling size or perhaps fledging survival was enhanced in nests where eggs were removed.
As described above, most male pied flycatchers tend to defend a single territory during the season. An exceptional few however, attempt to entertain polygynous relations with two females despite the costs that it entails to the secondary female. The male benefits, and so he tries to entice females onto his territory. Does the additional brood lead to a cost of reproduction that is measured in terms of either the males own survival, reproductive success the next season, or perhaps are his offspring affected?
If we are talking about male reproductive success, then we must consider the possibility that his own quality as mate might be affected from the point of view of mate attraction, likewise the quality of his offspring.
Gustafsson and his colleagues from Uppsala University in Sweden have been measuring the effects of increased brood size, and monogamy versus polygyny in the pied flycatcher. The have used Lack's classic manipulations of clutch size to get at the costs and consequences of parenting.
The males use the white patch as a badge of status. Males with a large patch appear to be more vigorous and drive off other males. They also appear to be more vigorous to females, because the males with the largest patches tend to be the polygynous males. Incidentally, the white patch is very bright in the UV spectrum.
Yes, males that return to breed that have serviced as father to the offspring from two females have a smaller patch size the following year. This suggest that the path is also an honet indicator of the male's physiological quality.
Males that were monogamous tend to return with a larger patch and also tend to be the males that enter into a polygynous relationship with females.
Secondary females are more likely to settle with a male which a large patch size because the patch advertises the males above average condition, and that the male might be a better parent than a male with a small patch.
Females are using a male's patch size as an honest indicator of his physiological condition and his net worth as a parent.
Male offspring from enlarged clutches tend to return with a very small patch in their first year of breeding compared to offspring which are from control clutches. Offspring from the reduced clutch size treatments tend to return with extra large patches at maturity.
These, results suggest that offspring quality is strongly affected by the costs of reproduction that parenting imposes on both male and female parents. Such costs can even lead to affects on the offsprings success in attracting mates.
Birds lay eggs one at a time, usually separated by several days. They need to fly, so having a full clutch of eggs yolked up is not all that viable. This leads to some eggs hatching earlier and some later. Such asymmetries in hatch date lead to profound fitness consequences.
Research on the cattle egert has led to the startling conclusion that the birds always lay two eggs, but only in exceptional cases will the second chick survive. In fact, the first laid egg usually hatches first and when the second laid egg hatches, the first chick will torment the chick and usually end up killing it or force it out of the nest.
The amazing thing about this behavior is that it is helped by some strage maternal provisioning. Female cattle egrets put extra testosterone in the the first laid egg which makes the first check more aggressive. This gives the check even more of an edge over its sib-mate and allows it to be a more effective "Cain" during the killing of its "Able".
The adaptive explanation that has been offered for such behaviors, is that parental cattle egrets rarely have enough resources to feed two chicks up to fledge. Under exceptional circumstances they might. But, most of the time, the let the offspring kill off its sib because that way at least one chick fledges.
It is not always the case that females give the first laid eggs extra testosterone. Hubert Schwabl has found that in birds that can raise large clutches, the female will give the last laid eggs extra testosterone to give them a fighting chance against their older sib mates. The testosterone helps the chicks at being better beggers and this allows them to catch up and be about the same size as their sibs despite the fact that they are not as old.


