
Speciation
An example of adaptive landscape in bacteria will be outlined in lecture.
Biological species concept -- "Species are groups of actually or potentially interbreeding populations, which are reproductively isolated from other such groups." Mayr 1942
Hybrid sterility definitely separates two groups as species though it is not the sole criterion
Reproductive isolation could however be due to behavioral or morphological
blocks to mating (i.e., hybrids are possible but unlikely given such blocks).
Barriers are not merely geographic, but have a biological origin
and arise during the process of speciation.
Isolating mechanisms:
A) Premating isolating mechanisms -- prevent union of gametes
-> zygote
B) Post-mating isolating mechanisms -- varying degrees of hybrid
sterility
And, we can have both A) premating and B) post-mating simultaneously
The biological species concept begins to break down in areas or at times
of speciation or incipient speciation (semispecies). For example
at hybrid zones, the somewhat differentiated populations are in the
process of speciation.
Genetic similarity drops as we change from the different levels
of differentiation (c.f., Wright's population subdivision). (overhead)
Populations - Subspecies - Semispecies - Sibling species - Non-sibling species -
Species -- NO GENE FLOW
Clearly genetic differentiation is occuring but it is not enough to define a species. It can vary from organism to organism and from mode of speciation to the next.
Migration is the movement of animals between what are referred to as populations of interbreeding animals. Rates of migration are usually expressed as the proportion of individuals in a population that disperse from one population to the next each generation. If migration between the region of contact between the two semispecies and the rest of the semispecies ranges is too high, then the two semispecies will become homogenized into one species even if such interbreeding results in lower fitness because of hybrid unfitness (Endler, 1977). If migration is low enough, then the two pure semispecies will eventually become differentiated into two species owing to the evolution of isolating mechanisms. Evolution of such isolating mechanisms is contingent upon the degree of genetic similarity between the two semispecies.
Figure 5.1. Hypothetical hybrid "unfitness" resulting from an incompatibility between genomes of the two parent species.
Large differences in genetic similarity between semispecies are
more likely to produce unfit hybrids than small differences in similarity.
If the two semispecies have many genetic loci that differ in the kind of
allele that they possess, then those alleles might not work very well together,
and more importantly the way two different genetic loci work together may
begin to breakdown. This is referred to as genome-wide genetic interactions.
The correct operation of one genetic locus (lets say an enzyme) may require
a particular form of the enzyme that is produced by a different locus (e.g.,
epistasis) (Whitlock et al 1993).
These enzymes are most likely to be compatible in their function if they
come from the same species. If they are from different species, they may
not integrate well with other loci in the organism and the hybrid individual
may die. Imagine such genetic interactions occurring at all possible loci
(e.g., 10,000 loci in Drosophila or perhaps 100,000 loci in humans). The
probability of a hybrid being more unfit has to do with how many loci in
each semispecies are homozygous for different alleles. Hybrid breakdown
occurs when many of these loci are fixed for different alleles in the two
subspecies and the production of offspring by the hybrids is not possible
because and F1 F1 produces too many incompatible gene by gene combinations.
MODES OF SPECIATION -- CLASSIFIED BY GEOGRAPHY.
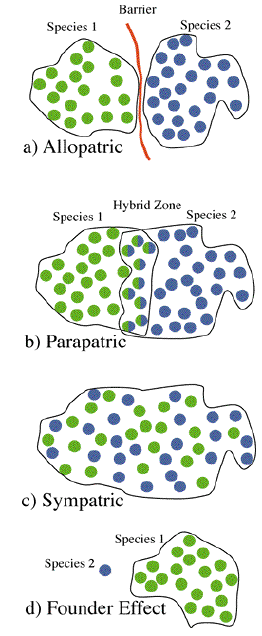 To understand the origin of species
differences it is necessary to consider how genetic differences arise between
species. Geographic subdivision decreases gene flow. Depending on the population
size, genetic drift can play a major role in promoting genetic differentiation.
If population size remains low for a long period of time it is possible
for genetic differences to accumulate between geographic areas by the random
process of genetic drift. Selection can promote speciation because genetic
changes can rapidly accumulate in one area where selection favors certain
behaviors, physiology, or morphology relative to another area where alternative
alleles are favored by natural selection. Finally, the amount of gene flow
which is usually described in terms of the migration rate is a key parameter
that influences the rapidity of speciation. By lowering gene flow, geographic
subdivision promotes the process of speciation (Endler, 1977). Ernst Mayr
(1942) has argued that some form of geographical subdivision is required
for speciation.
To understand the origin of species
differences it is necessary to consider how genetic differences arise between
species. Geographic subdivision decreases gene flow. Depending on the population
size, genetic drift can play a major role in promoting genetic differentiation.
If population size remains low for a long period of time it is possible
for genetic differences to accumulate between geographic areas by the random
process of genetic drift. Selection can promote speciation because genetic
changes can rapidly accumulate in one area where selection favors certain
behaviors, physiology, or morphology relative to another area where alternative
alleles are favored by natural selection. Finally, the amount of gene flow
which is usually described in terms of the migration rate is a key parameter
that influences the rapidity of speciation. By lowering gene flow, geographic
subdivision promotes the process of speciation (Endler, 1977). Ernst Mayr
(1942) has argued that some form of geographical subdivision is required
for speciation.
Figure Legend. Models of speciation based on geographical subdivision.
Models of speciation relate to the degree of geographical subdivision for
Allopatric --> Parapatric --> Sympatric which ranges extreme to none.
Founder effect speciation is an extreme form of subdivision in species formation
takes place in a small isolated population.
Founder Effect or Peripatric Speciation -- speciation occurs in an isolated population.These relate to the degree of geographical subdivision which ranges from:
Extreme <--> None
for the modes: Allopatric -- Allo-parapatric -- Parapatric -- Sympatric
Patterns -- Variation within a species is at the heart of the
matter.
CLINES:
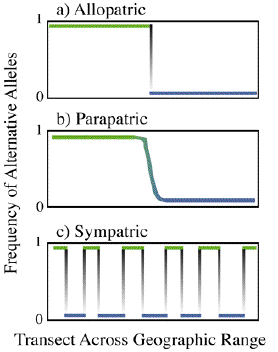
Big Question: Can we distinguish between primary and secondary
contact?
ALLOPATRIC SPECIATION
By isolation of a colony (e.g., island and mainland)
By division of range by extrinsic barrier or extinction of populations
By geographical distance.
Ring Species are a peculiar form of speciation that takes place
when the center of a species range is effectively unoccupied and thus there
is no gene flow. Adjacent races will interbreed, however the races at the
termini are so divergent, that there is no interbreeding (exist in sympatry).
Fig. 3, p 225 (of Futuyma) Thamnophis (garter snakes in California) (overhead)
Fig. 4-4, p. 65 (of Price) Sea Gulls and the Arctic Ocean
Hybrid zones -- Tend to show underdominance in fitness, that is
heterozygotes (hybrids) have lower fitness, and this is taken as evidence
of the hybrid zone being due to secondary contact.
Introgressive hybridization -- If there is any viability of the
hybrids, alleles may leak across and introgress across the zone. Some alleles
may spread quite far across the zone whereas others will be quite abrupt
and coincide with the hybrid zone.
PARAPATRIC SPECIATION
Clines -- may arise from environmental difference and selection.
Consider a simple population genetic model. Selection along a string of demes.
(From Figure 6, p. 162, chap VI, of Futuyma). (overhead).
In this model the form of the cline is primarily determined by selection.
A second model by Slatkin (1973) -- The width of the cline arising from
selection is given by s/where s
is the standard deviation of dispersal distance, and s is selection
against the wrong allele.
Why should the clines in a number of loci be concordant?
linkage - - alleles are "dragged" along (requires many such linkage groups)
epistasis - - fitness at one locus depends on genotype subject
to clinal selection
If the cline is primary (i.e., not from secondary contact), then there
is the possibility that parapatric speciation was responsible for the divergence.
However, parapatric speciation is greatly debated.
SYMPATRIC SPECIATION
One form is widely accepted -- the instantaneous mode in which
changes in ploidy lead to instantaneous reproductive isolation (e.g., in
plants). The examples from chapter 22 are used in lecture.
Most other theories involving a gradual change in sympatry are controversial.
One class of models maintains that there arise multiple-niche polymorphisms (heterozygotes inferior). Assortative mating among homozygotes is a key component of this theory. This minimally requires two loci to evolve simulatneously, and thus we establish strong linkage disequilibrium for the niche polymorphism loci and the assortative mating loci. More loci makes it unlikely.
Sympatric Speciation
The evolution of such divergence, in the face of gene flow is a tremendous challenge for the evolution of species by the process of sympatric speciation. In a classic population genetic analysis, Felsenstein (1981) showed that the process of sympatric speciation is a fairly unlikely event. He modeled this process in terms the evolution of two species attributes: 1) the phenotypic attributes that give rise to hybrid unfitness, and 2) the genes that give rise to mate discrimination. Generally, the genes for fitness, are not expected to be linked (either by pleiotropy or by physical linkage) to the genes that give rise to mate discrimination. This key fact places a tremendous genetic constraint on speciation by the process of reinforcement. Every generation segregation of chromosomes will lead to parental species that lack the correct discrimination alleles. Even if the genes fitness and mate discrimination are linked on the same chromosome, recombination during meiosis is a powerful enough force to dissolve away such associations and eliminate the possibility of sympatric speciation.- from the time of the modern synthesis up until very recently, the importance of sympatric speciation was greatly downplayed.
- the main reason for this was Ernst Mayr.
- Mayr championed the allopatric mode of speciation, and there is little doubt that Mayr is correct in recognizing the predominance of allopatric speciation in nature.
- however, Mayr was very adamant about allowing that even a small portion of species may be speciating by the sympatric mechanism.
- one of the major reasons why evolutionary biologists now consider the possibility that sympatric speciation is a more common mode of speciation than originally thought has been a result of the work of Guy Bush.
- Guy Bush was a recently graduated Ph.D. student at Harvard in 1966, when he gave his first paper at a scientific conference.
- he was nervous and had a right to be nervous.
- his Ph.D. work had pointed to the possibility that a new species of Rhagoletis fruit fly was forming right beside its parent population by the process of sympatric speciation.
- situated in the audience listening to his talk were Dobzhansky, Mayr and other leading figures of the modern synthesis.
- according to the story, after he finished his talk, there were a few moments of deafening silence from the audience.
- then Dobzhansky spoke - he said "That's very interesting, but I don't believe it. Sympatric speciation is like the measles. Everybody gets it, but they soon get over it."
- Bush never quite got over it - he pursued
the Rhagoletis story for the next 20 years, making it one of the
best known cases of sympatric speciation known to date.
Simple model:
2 loci involved H locus = "host selection"
locus
S locus = "larval survival" locus
- suppose a species of fruitfly exists that mates and lays eggs on apple trees.
suppose: H1 = prefer apple
S1 = survive on apple
- the species would be H1H1S1S1
- suppose a new allele emerged, H2 that caused a shift in host tree preference from apple to cherry.
- if survival of the larvae on cherry was
low, a new allele at the S locus, S2, might occur that greatly
increases survival on cherry but does not survive on apple.
H2 = prefer cherry
S2 = survive on cherry
- selection would thus favor the H2H2S2S2
genotype on cherry, which could lead to a new species with different host
preference.
- the crucial factor is what heterozygotes tend to do.
- however, these fruitflies have a strong tendency to remain and oviposit on the tree in which they emerged.
- this might explain in this example why speciation
might occur so readily.
GENETIC THEORIES OF SPECIATION -- Templeton's Cohesion
Species Concept this is a completely different
classification scheme from the Geographic modes of speciation. (Genes are
the focus not geography).
Problems with peak shift
Given saddle, peak -- the probability of peak shift is prop. to Ne. If Ne is small, the probability of peak shift is large, if Ne is large, the probability of peak shift is trivial.
FOUNDER EFFECT (PERIPATRIC SPECIATION) Mayr's Theory
Arguments against founder effect: Sampling effects are only important if inbreeding kicks in (the population remains bottlenecked for lots of generations.)
Arguments in favor: