Back to the Syllabus
7. Male Reproductive Strategies
Barry Sinervo and Yoni Brandt©1997
Index
Alternative Male Strategies
The Evolutionary Stable Strategy (ESS)
Alternative Male Morphs
Parental versus Satellite Behavior in Bluegill
Sunfish
Marine Isopods and Alpha, Beta, and Gamma Males
Equal Male Fitness versus Frequency Dependence:
Pattern versus Process Explanations for Morph Diversity
Aggression, Male Assessment, and ESS games
The Hawk-Dove and the Evolution of Lethal Fighting
The War of Attrition
Display of Intentions
Neighbor Recognition
Tit for Tat
Asymmetric versus Symmetric Contests and Badges
of Status
The physiological and behavioral basis of Resource
Holding Power
Deceptive Strategies
The Rock-paper-scissors game and the evolution
of alternative male strategies
Introduction to the rock-paper-scissors game
Female Behaviors towards Males
"Play-off" matrix and male behaviors
Alternative Male Strategies
In the introduction to Sexual Selection,
I described two ways of generating variance in mating success:
- female choice
- and male-male competition.
It is important to realize that these two avenues for selection are not
necessarily mutually exclusive. Traits that lead to female choice may also
be effective weapons during male-male competition. For the moment, we will
take up the idea of male-male competition and explore the link between female
choice and male contests next week.
The Evolutionary Stable Strategy (ESS)
In previous lectures we developed the idea of an evolutionary stable
strategy as a behavioral phenotype that cannot be invaded by a mutant
strategy. When a mutant arises in a population of individuals that display
the ESS phenotype, the mutant does not spread. By a mutant strategy,
I mean any behavioral phenotype that does things in a way different from
the ESS. Seems like a pretty broad definition. The ESS must be uninvadable
to any strategy that the scientist can conceive (...based on the biology
of the organism). If the ESS can be invaded, then you might expect to find
a diversity of behavioral types in the population.
A second condition for the ESS is that it should also be a strategy
that has a definite advantage when rare. For example, in the t-allele example of genic selection the t-allele
clearly has a tremendous advantage when rare and will increase in frequency
at the expense of the wild type allele. In the pure homozygous form all
males would be sterile and the population would go extinct. Therefore, the
t-allele cannot exist in the pure homozygous form and thus, is not an ESS.
The case of the t-allele introduces the concept of a mixed strategy. The
two alleles will exist in a stable frequency of t- and +-alleles and neither
will be eliminated from the population.
In general, when we consider ESS analyses we are likely to show that
many populations will exist in a mixed strategy state. We are not usually
trying to find an ESS which would consist of a pure strategy. How boring
life would be without a little diversity.
Why we are interested in differences in male reproductive phenotypes?
When Behavioral Ecologists go out into natural populations, they invariably
find that males have very different ways of trying to get mates. However,
in some populations there appears to be only one kind of male type. Trying
to find an explanation for this kind of among population variation is the
goal of behavioral ecology. Trying to explain the coexistence of a diverse
set of behavioral types is also the goal of these kinds of studies. How
can such diversity be maintained in the face of very strong selection favoring
one type over another?
Index
Alternative Male Types
What kinds of male types are you likely to encounter if you head out
into the wild? Well alternative males strategies are usually broken down
into the following kinds:
Aggressive males that tend to have many females and are called
polygynous.
Regular old male that is monogomous and usually has one mate
(the standard by which we measure male success).
Sneaker or Satellite males that are usually not very conspicuous,
but live around the edges of the more dominant males in the population.
The kinds of males present in the population also depend on the kind
of care that the species provides to its young. For example, male birds
tend to be monogamous and may also participate in the rearing of young.
However, in fish, the parental male types need not be monogamous, for a
parental male fish could entice many females to spawn in the nest
that he guards.
To understand Alternative Male Strategies a little better we will explore
two model systems for studying male reproductive success:
- Parental versus satellite males in the sunfish,
- The alpha, beta, and gamma male types in a
marine isopod.
These two examples contrast the control mechanisms
underlying the development of the alternative male strategies. In one case,
it has been argued that the male types are condition-dependent strategies
and environmental factors influence development. In the other case it has
been demonstrated that the types are genetically determined.
Index
Parental versus Satellite Behavior in Bluegill
Sunfish
Male Bluegill sunfish come in three different size morphs:
- A large territorial parental male that courts females, and then
defends a nest in which he rears eggs that the female oviposits,
- A medium sized satellite or sneaker male that mimics females,
interupts a courting territorial male and attempts to fertilize the female,
and
- A very small satellite that dives in between a mating territorial male
and female and squirts ejaculate in an attempt to fertilize the
female.
Bluegill sunfish have external fertilization and Mart Gross has calculated
the probability that a male of a given morphotype will be successful in
fertilizing a female from the amount of time he spends in the vicinty of
females. Based on these calculations, males of all three mating types have
approximately equal fitness.
An important component of these three behavioral strategies is the fact
that the male types mature at different ages. The territorial males mature
in six to seven years whereas the other two male types mature in two to
three years. In addition, the two smaller morphs do not have all the costs
associated with parental care strategy of the territorial male. The other
two morphs parasitize the parental effort of the territorial male.
Eggs do not survive unless they are cared for by a territorial male. In
terms of ESS, neither of the two smaller male morphs can exist in a pure
form, whereas the territorial male can successfully do so. However, the
two smaller morphs may successfully invade the large territorial male.
Gross has argued that such morphs are condition dependent tactics in
which all males have the capability to become any of the alternative types.
As a result, " developmental decisions" are made, based up the
condition of the male. Males that are in poorer condition become the smaller
male types and the males that are in the best condition adopt the territorial-holder
strategy. Such condition-dependent strategies are contrasted with
genetically-based control over morphotype development.
Index
Marine Isopods and Genetically-based Strategies
Marine isopods come in three dramatically different size morphs:
- a large alpha male with elaborate horns coming out the rear end
- a medium-sized beta male which is about the same size as a female
- and tiny gamma males that are much smaller than females or males.
What do these male types correspond to in terms of behavioral strategies?
The large alpha males will toss other males out of the sponge if they get
hold of them. Thus they rely on their strength and large size to toss out
other alphas -- the biggest alpha might have a better chance of tossing
out smaller alphas. The beta males, on the other hand, impersonate females,
and solicit courtship from an alpha! And finally, Shuster has described
gammas as little sperm bombs (personal communication). They lurk around
the sponge waiting for the opportunity to dive bomb a pair in copulo.
Shuster has also shown that there is a fairly simple genetic basis to
the three types of males with the traits being controlled by one locus with
three alternative alleles (the details need not concern us here). The genetic
control of the isopod mating system may be different from that seen
in Bluegill sunfish which are thought to be conditional strategies.
Shuster and Wade addressed the first issue regarding the stability of the
three kinds of males. In the long run, will one morph tend to dominate because
it has higher fitness?
The null hypothesis in this case is that the morphs have the same
fitness (male morphs might be expected to persist in the long run).
The alternative hypothesis is that one morph has higher fitness,
and would tend to increase in fitness (and perhaps in the long run become
the only male type present in the poulation).
Shuster and Wade found that they could not reject the null hypothesis,
they did not detect any statistically significant differences in the mean
number of females that each male type was likely to produce. No evidence
suggested any gross difference in the number of matings achieved by each
morph.
Does this mean that there is no sexual selection on isopods?
Well not really... Individual alpha, betas, and gammas did show variance
in mating success. Some aphas mating more successfully than others. There
may be strong selection among alphas to be a good alpha (e.g., strong fighter),
or for betas to be "good at being sneaky". Shuster and Wade's
study was also conducted over many generations in natural populations and
they computed mean success averaged over all of these generations reflecting
the long-term equilibrium -- all males persist in the population.
There may also be differences among the male types if one considers shorter
time scales, and smaller spatial scales.
For example, Shuster and Wade had evidence that fitness in ispopods was
also strongly dependent on who was in the "neighborhood". These
isopods live in sponges with little chambers. A large alpha may have other
alphas, betas and gammas present with him or the alpha may be alone. The
kinds of neighbors that the male has, influences how successful he is as
an alpha.
For example, an alpha that lives in a chamber with a single female and
a single beta is going to sire the offspring of the female. The beta will
not be successful against the alpha.
However, an alpha with lots of females is at a disadvantage with a single
beta. The beta is likely to sire 60% of the offspring. This could be due
to sneakiness of the beta males when the alpha has lots of females to keep
track of. The beta manages to copulate with at least enough females so that
he is the sire of most of the offspring.
When the fitness of a male depends on the frequency of the other mating
types, we refer to sexual selection as being frequency dependent.
It is the nature of frequency dependent selection that determines who
will win and who will loose in the long run, or if all male mating types
will be preserved. A key condition that would prevent a unique ESS (e.g.,
only one male type wins) would be an advantage to each strategy when
rare. This is the another way of stating that a rare mutant can always
invade the population. Consider a male morph that is declining in frequency
because one other type has an advantage. If it has an advantage when rare,
then it will be protected from elimination in the population. As it becomes
rare, the fitness of the rare morph increase. We will explore this idea
in greater detail using the example of the rock-paper-scissors game that
is played out in lizards.
Index
Equal Male Fitness versus Frequency Dependence:
Pattern versus Process Explanations for Male Diversity
Arguments about the existence of male strategies that are based on a
premise of equal fitness are inherently arguments about the pattern one
would expect to see in natural populations. Males should have equal fitness
if they are to be stable over the long-term.
In contrast, arguments about the frequency dependence of the male strategies
are arguments that describe the process of natural and sexual selection.
Such process-based explanations have more merit as they might be used to
predict the dyamics of evolutionary change.
Finally, the equal fitness arguments are counter to the way we traditionally
construct null versus alternative hypotheses. The null hypothesis is that
the male types have equal fitness and thus the morphs will persist in a
population. The alternative hypothesis is that the morphs do not have equal
fitness and one morph will ultimately pervail. We know that there are a
diversity of morphs so the goal is really to collect enough data to accept
the null hypothesis. This is contrary to the way we traditionally construct
null versus alternative hypotheses. We usually want to collect enough data
to reject the null hypothesis and accept the alternative hypothesis.
The process-based arguments avoid such statistical problems by identifying
the environmental conditions that leads to a change in fitness. In the case
of male strategies, it is the social environment that leads to a change
in fitness. As a morph becomes rarer because it has low fitness when competing
against the rare mutant, a point will be reached when the fitness of the
declining morph turns around. It becomes so rare that it begins to increase
in frequency. The advantage of a rare morphotype is a powerful mechanisms
for preserving variaton.
Index
Male Assessment and Male Strategies
ESS models have been used extensively in evolutionary theory (Maynard
Smith and Price 1972, Maynard Smith 1982, Maynard Smith and Parker 1976)
to account for various features of animal fighting behavior, including:
- displays and communication in agonistic encounters,
- badges of status,
- lethal fighting,
- the effects of various asymmetries among contestants and their strategic
decisions.
All of these factors are believed to be involved in assessment of rivals,
and have been incorporated into various models. Evolutionary game theory's
early treatments of contest behavior was completely devoid of assessment
-- the act of acquiring information regarding your rival's abilities. Symmetric
contests were modeled, where contestants are identical in all but strategic
properties (i.e. choice of behavior). In effect, there is nothing to assess
in this type of contest, especially since conditional strategies are not
included in the strategy set. These early models include the Hawk-Dove game
and the War of Attrition. More advanced models consider information transfer
between contestants and male assessment.
The Hawk-Dove Game and The Evolution of Lethal
Fighting
The Hawk-Dove game was designed in response to a ubiquitous phenomenon
in animal conflicts: although contests are frequently vigorous, lethal
fighting is rather rare.
How can this be explained in individual selection terms?
What is the evolutionary value of restraint?
The Hawk-Dove contests considers two strategies:
Hawk, which fights at the drop of a hat, always fights, even against
other hawks.
The second strategy, Dove, backs down as soon as the contest escalates
to a fight.
The theory for the hawk-dove game shows that Hawk is an ESS that Dove
cannot invade, yet Hawk can always invade Dove. However, if the cost of
fighting is too high for Hawks, then Doves can invade because Hawks do each
other in during their contests.
Additional references
Maynard Smith and Price. 1972. The logic of animal conflict. Nature 246:
15-18.
Index
The War of Attrition
The Hawk-Dove game shows that the advantage of fighting behavior can
be offset by the cost of fighting, and that a cost benefit analysis can
show that when resource value is low compared to cost of fighting, restraint
is adaptive (actually this will be a mixed ESS).
Another simple model is the Symmetric War of Attrition , also proposed
by Maynard Smith. This is also known as "the waiting game" because
the winner of the contest, the individual that gains the resource, is the
individual that is prepared to persist longer. This is a situation where
there are an infinite number of possible strategies (persistence times),
a continuous strategy set. The cost of persisting is assumed to increase
monotonically through time. When one individual gives up, the other contestant
gets all of the resource, and both individuals incur the cost of persistence
to that point in time. Obviously each individual would do better by persisting
longer than its opponents, and thus no pure persistence level would be evolutionarily
stable. Mixed strategies allow an equilibrium: an unpredictable persistence
time would be evolutionarily stable. At the ESS, the fitness gain for each
persistence time is equal. This is a game where no information is exchanged,
since you can not learn how much longer your opponent will persist by observing
how long your opponent has already stayed. Given this ESS, no assessment
can occur here. This theoretical result was used to suggest that animal
contests in general should not involve any displays of intent (in this model:
persistence time), because such displays could be exploited by opponents.
Of course, animal contests are usually NOT symmetrical !
Contests usually involve displays.
What is the evolutionary value of displays in animal contests?
Males can learn and advertize asymmetries.
Index
Displays of Intentions
The prediction that animals in contests should not
reveal their intentions was derived from the symmetrical war of attrition,
although this prediction was valid only under very restrictive conditions.
In particular this was a symmetrical game in which the combatants were equally
matched in ability and resource value. Revealing intent was not evolutionarily
stable, since it could be exploited by cheaters using 'cheap talk'. If a
cheater could detect intent, then it could try to bluff it's way out of
the contest by exploiting the information to it's advantage.
Contrary to this prediction, empirical evidence quickly accumulated that
showed that animals in fact do often vary their choice of display in
a manner that is predictive of future behavior, and several models were
constructed to explain why such behavior could be evolutionarily stable
when they should not reveal much information. Enquist proposed a model ,
nicknamed the 'risk-right' model. This model suggested that cheating is
precluded because increasingly effective threat displays are increasingly
costly in terms of risk of retaliation. Thus only individuals that are actually
prepared to pay a high probability of retaliation can afford to use highly
effective displays. This model predicted that display types that are increasingly
effective in winning disputes are also increasingly likely to cause retaliation.
Subsequent empirical work has supported this prediction.
Another model by van Rhijn and Vodegel, suggests that cheating is precluded
because individual recognition will allow the detection of cheats,
and select against cheating. Many animals display some form of neighbor
recognition.
Index
Neighbor Recognition
The basic experimental paradigm for testing for such rudimentary cognitive
abilities involved in neighbor recognition is to use a playback
experiment in birds in which a tape recording of the neighboring male's
song is played at the territory boundary (Brooks and Falls, 1975; Wiley
1977, Godard, 1991). The response of the target male to the neighbors song
is usually less extreme than the targets response to a strange male's song.
Why is this the case? The Dear Enemy Hypothesis maintains that once
males have established territory boundaries it is a waste of the male's
time to continue in escalated conflicts with the neighbor. The two exist
in a status quo in which they are still assessing one another, but presumably
neither is likely to act aggressively because the other male has a home
field advantage. Each male also has access to females and food and thus
the motivation to "cross the line drawn in the sand" is
low for both males. We can express these ideas in terms of the marginal
value theorem with the added twist of risk involved in conflict. The marginal
gains that are derived from increasing territory size are not worth the
risks of an all out battle. However, if the bird hears a stranger at the
boundary, the male must act to turn away the intruder. A targeted male in
a playback experiment is much more agitated when he hears a strange male's
song coming out of the speaker than if he hears a neighbor. Birds even have
the capability to remember a neighbor's song across reproductive seasons
(Godard, 1991).
Such neighbor-stranger experiments have also been performed on lizards,
but in this case a challenger (stranger or neighbor) is tethered at some
point in the targets territory, and the reaction of the target is videotaped.
Stan Fox has found that the target is more agitated in response to strangers
than to his dear-enemy neighbor.
Additional references
Falls, B. and Brooks, R.J. 1975. Individual recognition by song in white-throated
sparrows. I. Discrimination of songs of neighbors and strangers. Can. J.
Zool. 53: 1412-1420.
Falls, B. and Brooks, R.J. 1975. Individual recognition by song in white-throated
sparrows. II Effects of location. Can. J. Zool. 53: 1412-1420.
Godard, R. 1991. Long-term memory of individual neighbors in a migratory
songbird. Nature 350: 228-229.
Fox, S.F. and Baird, T.A. 1992. The dear enemy phenomenon in the collared
lizard Crotaphytus collaris, with a cautionary note on experimental methodology.
Anim. Behav. 44 780-782.
Index
Tit for Tat Among Neighbors
In the dear enemy situation it is thought the the neighbors have some
sort of status quo in effect. You don't whack my back, I won't whack yours.
What happens when one neighbor breaks such conventions (a non-aggression
pact if you will), and invades the other males's territory? Rene Godard
used the power of the playback experiment to test whether such "defecting"
neighbors are treated differently after the incursion. She played the songs
of neighbors on the territory boundary where both the nieghbor (N) and strangers
(S) should reside. In addition she also played the song of the neighbor
on the other side of the territory (XN), a place where the neighbor had
no "right" to be. After such wrong-place incursions, the territory
holder was usually much more aggressive at the boundary of the neighbor
than if the playback occured in the proper place. The targeted bird went
to the neighbors border and sang more as a kind of tit for tat response
to the XN intrusion. The intrusion triggers much more aggression in the
targeted male towards the perceived neighbor that intrudes.
Additional references
Godard, R. 1993. Tit for Tat among neighboring hooded warblers. Behav.
Ecolo. Sociobiol 33: 45-50.
Index
Asymmetric versus Symmetric Contests and
Badges of Status
Of course most animal contests involve asymmetries. A lot of attention
has been directed at the effects of asymmetries on the strategic choices
of animals. The most recent of these models are characterized by explicit
treatment of acquisition of information about the opponent. War of attrition
models first introduce asymmetry into contest behavior, of three types
- The simplest of the asymmetries can be thought
of as a home field advantage. It is invariably the case that a territory
holder has an advantage over intruders. <Experiment on Butterflies by
Gilbert>
- The second kind of asymmetry is one in which one constestant has
more to gain or loose from the battle and will thus be motivated to continue
a prolonged contest. A classic case involves a territory holder versus
an intruder. The territory holder has a lot invested in the territory.
He has knowledge of safe retreats, the dear enemy relationships that have
been constructed, knowledge of resource. All of this information makes
the territory more valuable to the territory holder than to the intruder.
To reconstruct all this information in a new territory would require a
lot of effort.
- The most interesting kind of asymmetry is one in which individuals
vary in their physical ability to hold onto the territory or resource --
some are more capable fighters. This is termed Resource Holding Potential.
The sequential assessment game models the acquisition of information
within a contest and injects explicit decision mechanism into models of
contest behavior. As the contest unfolds, each contestant accumulates information
about the opponent. Once enough data has been collected, a male may decide
to end the contest and flee, or escalate to the next stage of the contest
to acquire more and more information. Different displays and levels of escalation
measure different attributes associated with fighting ability. Diminishing
returns of increasing sample size within a single class of displays leads
to escalating to the next display in which more information about the opponent
might be gained. The aim of assessment strategies is to determine the Resource
Holding Power or Potential of the opponent.
In asymmetric contests a male mating type
which is stronger than the others would be expected to "advertise"
this skill by some sort of badge of status. By walking around and
displaying this badge of status, a very dominant individual could avoid
most contests because other males would be unlikely to challenge such a
powerful or skillful male. By avoiding unnecessary contests, the dominant
individual could focus on contests with other more equally matched individuals
that are likely to lead to a War of Attrition. Some advertising may be necessary
on the part of the most dominant individuals even though the simple war
of attrition models suggest otherwise, because the dominant can avoid many
contests.
Index
The Physiological and Morphological Basis
of Resource Holding Power
Resource Holding Power and associated Badges of Status have tremendous
advantages in male assessment games, but those strategies are usually not
cost free. In vertebrates, male secondary sexual characteristics such as
large size, or elaborate ornaments and armaments are usually related to
levels of circulating testosterone. More and more researchers are beginning
to study the negative effect that such steroids have on other aspects of
the organisms life history. For example, Marler and Moore (cited in Alcock)
have shown that males implanted with T have reduced survival compared to
controls in field trials. Such survival costs can be ameliorated by supplementing
the T-implanted males with food. Thus, while badges of status are useful
in information transfer that can avoid costly conflicts, such badges have
additional life history trade-offs that counterbalance the fitness gains
that arise from conflict resolution.
Classic examples of communication of Resource Holding Power include "roaring"
in red deer (Clutton-Brock and Albon, 1978). In playback experiments, male
stags that are presented with a red deer recording that plays for an unusually
long time are more intimidated than males presented with a normal bout of
roaring. They interpret the length of time that a male deer can roar as
an index of Resource Holding Power, if better condition is required to be
able to roar for a long period of times.
Another example of RHP communication involves the croak of toads. Males
that are larger possess a deeper croak and male size is presumably related
to RHP. Again playback experiments in which the frequency of the croak is
manipulated can be used to alter the behavior of the target male to be more
"intimidated".
Additional references
Clutton-Brock, T. H. and S. D. Albon. 1979. The roaring of red deer and
the evolution of honest advertisement. Behaviour. 69: 145-170.
Index
Deceptive Strategies
If a mating system evolves individuals with badges of status which advertise
their prowess, the situation is ripe for an interesting strategy to evolve
to exploit this information in some way. It is not necessarily possible
to evolve a better strategy which would entail additonal costs assoicated
with the development of more "firepower". The individuals with
a badge of status may have some weakness, perhaps in the perceptual system
that could be exploited. The evolution of female mimicry in males is undoubtedly
related to the exploitation of the information embodied in a badge of status,
and an ensuing weaknesses that strategy entails. Rather than fight, the
sneaker uses a deceptive strategy. We will explore this strategy in detail
in the rock-paper-scissors game
Index
The Rock-paper-scissors Game of Alternative
Male Strategies: A Case Study
 Amuse yourself
before lecture by exploring lizard land. Clicking on the links located below
will take you to the virtual rock paper scissors game. I will go over the
material in the following pages as a summary of the ideas presented on alternative
male strategies.
Amuse yourself
before lecture by exploring lizard land. Clicking on the links located below
will take you to the virtual rock paper scissors game. I will go over the
material in the following pages as a summary of the ideas presented on alternative
male strategies.
See Barry Morph into a Lizard.
Introduction to the rock-paper-scissors game
 An unusual game is being played out in the Coast Range
of California. Three alternative male strategies are locked in an ecological
"perpetual motion machine" from which there appears little escape.
As in the rock-paper-scissors game where rock beats scissors, paper beats
rock, and scissors beats paper, three morphs of lizards cycle from the ultra-dominant
polygynous orange-throated males, which best the more monogamous mate gaurding
blues; the oranges are in turn bested by the sneaker strategy of yellow-throated
males, and the sneaker strategy of yellows is in turn bested by the mate
guarding strategy of blue-throated males. Each strategy in this game has
a strength and a weakness, and there is the evolutionary rub that keeps
the wheels spinning. See B. Sinervo and C. M. Livley (1996) Nature 340:240-243.
An unusual game is being played out in the Coast Range
of California. Three alternative male strategies are locked in an ecological
"perpetual motion machine" from which there appears little escape.
As in the rock-paper-scissors game where rock beats scissors, paper beats
rock, and scissors beats paper, three morphs of lizards cycle from the ultra-dominant
polygynous orange-throated males, which best the more monogamous mate gaurding
blues; the oranges are in turn bested by the sneaker strategy of yellow-throated
males, and the sneaker strategy of yellows is in turn bested by the mate
guarding strategy of blue-throated males. Each strategy in this game has
a strength and a weakness, and there is the evolutionary rub that keeps
the wheels spinning. See B. Sinervo and C. M. Livley (1996) Nature 340:240-243.

|

|

|
| The Colors: Photos of the three flavors (orange, blue, and yellow) of male
side-blotched lizards, shown in profile and from the bottom of the throat. |
Copulation: These males spend most of their time trying to gain access to
females. Warning: the following image shows lizards engaged in copulation...click
at your own risk. |
Territory map: Orange and blue males actively defend a territory. Yellow
males tend to cluster around the territories of orange males, probably because
they are more successful in sneaking copulations from the females on an
orange male's territory compared to sneaking copulations from blue males. |
Access to the Video Library (also see matrix below)
Yellow-throated Sneaker Male Behavior
Yellow males are "sneakers" in that they mimic the throat color
of receptive females, and yellows also
mimic female behavior. When a yellow male meets a dominant male, he
pretends he is a female -- a female that is not interested in the act. The
head bobs ("vibrations") involved in actual female
rejection behaviors are of very high frequency compared to the low frequency
aggressive challenge displays shown by aggressive males. In many cases,
females will nip at the male and drive him off. By co-opting the female
rejection display, yellow males use a dishonest signal to fool some territory
holding males. The ruse of yellows works only on orange-throated males.
Blue-Throated Male Behavior
Blues are not fooled by yellows. Blue
males root out yellow males that enter the territory of the blue male.
Blue males are a little more circumspect when they engage another blue male
during territory contests. Blue males spend a lot of time challenging and
displaying, presumably allowing males to assess one another. Attack may
or may not follow as blue males very often back down against other blue
males. Indeed, Neighboring males use
a series of bobs to communicate their identity, and the neighbors usually
part without battle.
Orange-Throated Male Behavior
Orange males are ultradominant and very aggressive owing to high levels
of testosterone, and attack intruding blue males that typically have more
modest levels of testosterone. Attacks
by orange males on blue males do not involve the ritualized head bobs
that are seen when blue males engage blue
males -- orange males just attack with little advanced warning. When
an orange male encounters a more equally matched orange male, they are both
a little more circumspect, and they will not necessarily attack one another
(as is seen when blue meets blue).
Summary of the rock-paper-scissors game
Thus, each strategy has a strength and a weakness and there are strong
assymetries in contests between morphs. Trespassing yellows, with their
female mimcry, can fool oranges. However, trespassing yellows are hunted
down by blue males and attacked. While oranges with their high testosterone
and high stamina can handily defeat blues, they are susceptible to the charms
of yellows. In contrast, contests between like morphs (e.g., blue vs blue,
orange vs orange or yellow vs yellow) are usually more symmetric.
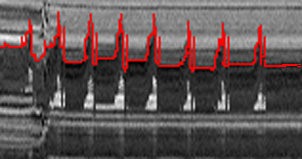 Analysis of Head-Bob Patterns
Analysis of Head-Bob Patterns
The bobs can easily be analyzed by digitizing images and "slicing"
through the video frames to get the outline of the lizard head bobbing up
and down through time. This has been highlighted in red. The male in this
sequence is participating in the "challenge" display. Yoni Brandt
and I are busily analyzing such videos.
Video Library of Male and Female Behaviors
in Side-blotched Lizards
 Film Credits:
Film Credits:
- Yoni Brandt
- Donald B. Miles
- Tosha Comendant
- Barry Sinervo
Outcomes of meetings between a male and a
female:
Pre-receptive |
Receptive |
 |
 |
Post Copulation |
Aggression |
 |
 |
Outcomes of morph encounters for the 9 possible
pairwise meetings:
Back to the Syllabus











