1. Outline
three hypotheses to explain why animals disperse from their natal site. What
are the levels of selection that are used in each hypothesis? (you must give
three biological examples, i.e., one of each hypothesis).
- inbreeding avoidance, such as in the
example that explains why males disperse further than females, which
implies that there is actually an advantage for one sex in kin
cooperation. Males gain no such advantage perhaps and thus disperse
(examples include lions, ground squirrels).
- Kin competition, if progeny compete
intensely, this favors a kin altruistic act on the part of one kin that
disperses, while the other remains philopatric, Side-blotch lizards,
(though a variation on the example in 1 could be used here, provided it is
a different species).
- Extinction probability due to
habitat loss favors the evolution of a dispersive form, from the viewpoint
of species selection.
Level used in
each: 1) individual and kin selection, 2) kin selection, 3) species (or group
selection).
2.
Describe the evolutionary scenario that leads from palatable to unpalatable
aposematic forms in terms of a learning experiment on Great Tits in a novel
world.
The
critical issue in this answer is to set up the experiment with obvious versus
cryptic forms. The obvious forms are defended with a noxious compound
(Chloroquinone), while the cryptic is undefended. The prey are of course loaded
with a treat (that is either yucky [with Chloroquinone] or tasty [without
Chloroquinone].
Abstract
geometrical symbols are attached to the “wings”, and these are either cryptic
or obvious relative to symbols that are randomly printed on paper that is
distributed on the floor of the aviary. [a figure with these salient features
will suffice for my lengthy explanation].
Then,
the obvious defended form is released in aggregations or it is released
solitary, cryptic forms are always solitary.
This
experiment tests Fisher’s (1930) classic idea that aggregations allow for kin
benefit to generate a learning effect in the predator (single-trial learning)
in which an aggregated defended form gains an advantage against a cryptic form,
or solitary defended form.
3. Compare and contrast acquisition of song in cowbirds and viduine
finches. Which group has higher rates of speciation and why? Which group can
parasitize the most species and why?
In classic song
learning a bird learns its song from its father (both female and male progeny),
and then sexual imprinting causes females to prefer such songs as adult
females, or sing such songs as adult males.
Viduine finches
are no different from classic songbirds, except that sexual imprinting is
transferred over to host searching behavior. It is still used in mate choice
(within species), but other signals are required (species specific morphology,
behaviors, besides song) that elicit female preference and loordosis. This
cultural loop, couples the good songs of parents (that found the nest), to good
songs in males that mimic the host, to good song recognition in females during
both mate search and host search in a culturally selected runaway, generating
phenomenal rates of speciation.
In cowbirds,
song is not learned but is innate and species typical. Female birds might still
learn the song of their host, but some species of cowbirds actually are
generalists, and can parasitize up to 250 different hosts. However, song itself
does not evolve by cultural evolution, rather evolves by standard genetic
processes. Males, during the song rehearsal period, refine their song during
interactions with females, a classic audience effect.
4. What is
a maternal effect? How is this different from genetic alteration of phenotype?
What are the advantages of a system in which the female can alter offspring via
a maternal effect, compared to hard-wired genetic differences?
A maternal
effect reflects some non-genetic effect that is given from mother to progeny
that alters the progeny in some way. No genes are passed to progeny to
specifically cause the alteration in phenotype, and often other substances like
hormones are passed, that cause the alteration.
There may in
fact be genes passed to progeny, which even govern this process, but these are
only activated in females, when they become reproductive, in the standard
Menedelian transmission route.
The advantage of
such a system is that a female can alter the progeny in a way that enhances
progeny survival or reproductive success. The female is essentially predicting
something about the offspring environment, which she has information on, but
the progeny may not.
Genes cannot
accomplish this kind of exquisite manipulation and fine-tuning of progeny
genotype.
For example,
adding yolk hormones to eggs in a bird that has asynchronous egg laying, and
biasing levels of testosterone to later hatched eggs, effectively is predicting
the harsher competitive environment faced by late hatching chicks.
Either the
example or the explanation of prediction will suffice.
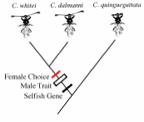
5. a) Does the example of stalk-eyed flies refute the idea of sensory bias
(need a phylogeny)? If it refutes sensory bias is it an example of indicator
sexual selection, runaway process or is it just a random pattern? Provide
evidence for your answer.
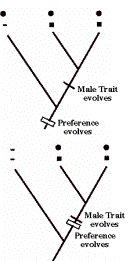
Sensory bias has
the female preference evolve prior to the outgroup, and the male trait evolve
after the split of the outgroup (top panel). A phylogeny that refutes sensory
bias would have female preference and the male trait evolve in concert (such as
in runaway, the bottom panel), or (optional) female preference evolve after the
male trait.
In stalk eyed
flies, the male trait female preference, and good genes for restoration of sex
ratio distortion appear to all evolve at the same time. The existence of good
genes implies a more complex model of sexual selection compared to runaway, and
the concerted evolution of all three clearly refutes sensory bias.
6. What is the prisoner's dilemma? A payoff matrix or a discussion of the rewards and punishments involved is essential (if you want you can make the payoff matrix apply to the following example). Describe one biological example (non-human) of tit-for-tat as a solution to the iterated PD.
|
|
Player B |
||
|
Player
A |
|
Cooperation |
Defection |
|
Cooperation |
-1 |
-10 |
|
|
Defect |
0 |
-9 |
|
Let us consider
all of the payoffs, where player A Defects given cooperation by B is termed
D|C:
D|C > C|C >
D|D > C|D
- D|C is referred to as the temptation to defect.
- C|C is referred to as mutual cooperation
- D|D is the punishment for mutual defection
- C|D is the sucker's payoff
Both players should defect (rationale), but the best payoff is actually cooperate.
Hypothetical payoff matrix for territorial hooded
warblers, Wilsonia citrina,
engaged in a dear enemy relationship with defection. This hypothetical payoff
matrix satisfies the conditions for the prisoner’s dilemma in that:
Temptation> Cooperation > Mutual Defection >
Sucker's Payoff.
Let us consider
the pay off matrix:
|
|
Neighbor |
||
|
Focal
Male |
|
Cooperation |
Defection |
|
Cooperation |
Cooperation-save energy by avoiding confrontations |
Sucker's
payoff looses
territorial resources
or females |
|
|
Defection |
Temptation to gains resources and females and keeps its own |
Mutual
defection --
come up even |
|
There are in fact
three play movements that underlie the Tit-for-Tat strategy:
Nice in which both players cooperate on the first move of
the game,
Retaliatory in which a player defects if an individual defected
on the prior move, and
Forgiving in which a player cooperates with a past defector
that now has chosen to cooperate.
Warblers do these behaviors in neighbor, stranger, neighbor-in-the-wrong-place experiments.
The second part of this example, would suffice for the entire answer.