Back to 4. Levels of Selection
Side Box 4.3: Inbreeding
Inbreeding can be defined in many ways, but we generally refer to the
simplest definition which has to do with matings among related individuals.
A more precise definition is to express inbreeding as the probability
of the two alleles on complementary chromosomes being identical by descent.
If these two copies of the allele (one from mother and one from father)
are identical by descent, then they must have arisen in the not-so-remote
past from a single strand of DNA.
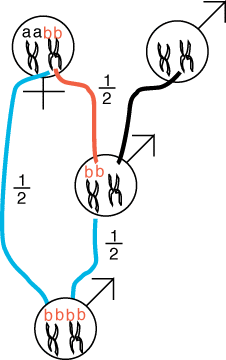 Genes that have alleles identical
by descent is can easily be generated from consanguinous matings,
or a mating among close relatives. Consider progeny fertilized from a mating
between a mother and son. We can track one allele, b,
from mother to son and from the mating of mother and son which we will call
an oedipal mating. Each chromosome is represented by two sister chromatids.
Allele b is located on one of the mother's
two chromosome pairs.
Genes that have alleles identical
by descent is can easily be generated from consanguinous matings,
or a mating among close relatives. Consider progeny fertilized from a mating
between a mother and son. We can track one allele, b,
from mother to son and from the mating of mother and son which we will call
an oedipal mating. Each chromosome is represented by two sister chromatids.
Allele b is located on one of the mother's
two chromosome pairs.
The son may receive one copy of allele b
with probability of 1/2 (denoted by the red
line). The blue lines denote the 1/2 probability
that the mother will contribute the b to a
progeny sired by her son, who also contributes allele b
with probability 1/2 via the second blue line.
Because all events are mutually independent, we multiply all of the probabilities
to get the net probability that a child produced by an oedipal mating possesses
two bb alleles:
1/2 * 1/2 * 1/2 = 1/8 (eqn 1).
Note that, the "a" copy could just as easily been transmitted
to the son in the first place so the total probability that the alleles
in the oedipal child are identical by descent can be achieved two ways,
either with allele "a" or with allele b
and the probability of inbreeding from an oedipal mating is:
2 * 1/2 * 1/2 * 1/2 = 1/4 (eqn 2).
Of course, there is a 3/4 probability that the oedipal child is not identical
by descent. Let us consider a few special cases to assess the fitness consequences
of such inbreeding.
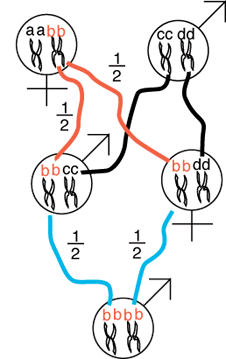 As a second example, let
us consider the possibility of a sib-sib mating. We now have to keep track
of the fathers alleles because a grandchild can become inbred from one of
the mom's alleles or one of the the father's alleles. Again, we will track
one allele, bb, from mother to son and from
the mother to daughter. At the end we will then multiply the probability
of a single allele being identical by descent by four (all possible alleles
that might be identical by descent) to determine the overall probability
that any allele is identical by descent.
As a second example, let
us consider the possibility of a sib-sib mating. We now have to keep track
of the fathers alleles because a grandchild can become inbred from one of
the mom's alleles or one of the the father's alleles. Again, we will track
one allele, bb, from mother to son and from
the mother to daughter. At the end we will then multiply the probability
of a single allele being identical by descent by four (all possible alleles
that might be identical by descent) to determine the overall probability
that any allele is identical by descent.
The mother gives the b allele to her son
with probability of 1/2 and likewise the daughter receives the b
allele with probability of 1/2 (both denoted by the red
lines). Given that the daughter received the b
allele, the daughter passes the allele on to her own progeny with propbability
1/2 (blue line). Likewise, given that the son
has the b allele he passes it on to his progeny
with probability of 1/2. The probability that the grandchild received two
copies of the b allele is given by:
1/2 * 1/2 * 1/2 * 1/2 = 1/16.
Finally, we realize that the grandchild could just as well have received
the a, b, c, or d alleles by the same routes with the same probabilities
so the probability of any two alleles at a single locus in the same individual
being identical by descent is given by:
4 * 1/2 * 1/2 * 1/2 * 1/2 = 1/4.
The same logic can be used to compute the probability of two cousins
yielding a child that has genes identical by descent, though the length
of the paths are a little bit longer. Indeed, the logic used above can be
used to compute the probability of inbreeding or identity by descent
for any consanguinous mating. The inbreeding in any set of pedigrees
can be computed from similar path diagrams that chart the genealogical
relationships among individuals.